Lunchtime on a late summer day 20,000 years ago
in the southwestern part of what is now France: A
small group of boys have been playing since midmorning, exploring the caves that are common in
their region, looking for old stone tools that have
been left behind by hunting parties. They are starting to get
hungry. They do not head back to their village for food: The
morning and evening meals will be provided by their parents and other adults in the tribe, but they are on their own
between those two meals.
At this time of year, the boys do not mind foraging on
their own. The summer has been rainy and warm, and a
large variety of nuts, berries, and seeds are beginning to
ripen. Because the summer growing season has been a
good one, small game such as rabbits and squirrels are well
fed and will make a good meal if the boys can manage to
catch one. They spend an hour or two moving from site
to site where food can be found, covering a couple of miles
in the process. They see a rabbit and spend 20 minutes
very quietly trying to sneak up on it before realizing that it
is no longer in the area. Even without the rabbit, they are
all happy with the amount of food they managed to find
during their midday forage. In mid-afternoon, they stop by a
stream for a rest, and then one by one they fall asleep.
Lunchtime on a late summer day in the early twentyfirst century, at a middle school in the United States:
A large group of children line up in the cafeteria to get their
lunch. They have spent the morning behind desks, doing
their school work. They have had one short recess, but
they will not have another during the afternoon. They have
a physical education class only once a week because budget cutbacks have meant that their school can afford only
one gym teacher for more than 1,200 students.
As the children pass through the cafeteria line, most of
them ignore the fruit, vegetables, and whole-wheat breads.
Instead, they choose foods high in fat, salt, and sugar:
chicken nuggets, fries, and cake. The children do not drink
the low-fat milk provided but instead favor sweet sodas
and fruit-flavored drinks. After they sit down, the children
have 15 minutes to finish their meals. Most of them would
say that they really like the food the cafeteria gives them.
When they are finished, they return to their classrooms for
more instruction.
At first glance, children in developed countries in the early twenty-first century are much
healthier than their counterparts who lived 20,000 years ago. They are bigger and more
physically mature for their age, and unlike their Paleolithic ancestors, they can reasonably
expect to live well into their 70s. They have been vaccinated against several potentially lifethreatening viral illnesses, and they need not worry that a small cut, a minor broken bone,
or a toothache will turn into a fatal bacterial infection. They are blissfully free of parasites.
On the other hand, a child from 20,000 years ago might have grown up more slowly
than a contemporary child, but upon reaching adulthood he would have had a strong,
lean body, with much more muscle than fat. He would not have spent a lifetime consuming more calories than he expended. If he were lucky enough to avoid infectious disease,
injury, and famine, in his middle and old age he would have been less likely to suffer from heart disease, high blood pressure, diabetes, and even some kinds of cancer than
would an adult living today.
Health and illness are fundamental parts of the human experience. The individual experience of illness is produced by many factors. Illness is a product of our
genes and culture, our environment and evolution, the economic and educational
systems we live under, and the things we eat. When we compare how people live now
to how they lived 20,000 years ago, it is apparent that it is difficult to define a healthful environment. Is it the quantity of life (years lived) or the quality that matters
most? Are we healthier living as our ancestors did, even though we cannot re-create
those past environments, or should we rejoice in the abundance and comfort that a
steady food supply and modern technology provide us?
In this chapter, we will look at many aspects of human health from both biocultural and evolutionary perspectives, and we will see how what we can learn from the
skeleton about life, health, and disease is used by forensic anthropologists in recent
criminal cases. We will see how health relates to growth, development, and aging.
We will consider infectious disease and the problems associated with evolving biological solutions to infectious agents that can also evolve. We look at the interaction
between diet and disease and the enormous changes in our diet since the advent of
modern agriculture. Finally, we will see how the skeleton retains clues about our lives
and deaths that can be used to solve criminal cases.
Biomedical Anthropology and Biocultural Perspective
Biomedical anthropology is the subfield of biological anthropology concerned with
issues of health and illness. Biomedical anthropologists bring the traditional interests
of biological anthropology—evolution, human variation, genetics—to the study of
medically related phenomena. Like medicine, biomedical anthropology is a biological
science, which relies on empiricism and hypothesis testing and, when possible, experimental research to further the understanding of human disease and illness. Biomedical anthropology is also like cultural medical anthropology in its comparative outlook,
and its attempt to understand illness in the context of specific cultural environments.
A central concept of biomedical anthropology is adaptation. As we have discussed in previous chapters, an adaptation is a feature or behavior that serves over
the long term to enhance fitness in an evolutionary sense. But we can also look at
adaptation in the short term; this is known as adaptability (Chapter 6). A basic question biomedical anthropologists try to answer is: To what extent is adaptability itself
an adaptation? For example, the life history stages that all people go through have
been shaped by natural selection, but our biology must be flexible enough to cope
with the different environmental challenges we will face over a lifetime.
In addition to an adaptation-based evolutionary approach, many biomedical anthropologists look at health from a biocultural perspective. The biocultural approach
recognizes that when we are looking at something as complex as human illness, both
biological and cultural variables offer important insights. The biocultural view recognizes that human behavior is shaped by both our evolutionary and our cultural
histories and that, just as human biology does, our behavior influences the expression of disease at both the individual and population levels (Wiley 2004; Wiley and
Allen, 2009).
An example of an illness that can be understood only in light of both biology and
culture is anorexia nervosa, a kind of self-starvation in which a person fails to maintain
a minimal normal body weight, is intensely afraid of gaining weight, and exhibits disturbances in the perception of his or her body shape or size (Figure 15.1) (American
Psychiatric Association, 1994). The anorexic person fights weight gain by not eating, purging (vomiting) after eating, or exercising excessively. The prevalence rate
for anorexia is about 0.5%–1.0% among teenaged and young women; about 90% of
all sufferers are female. Anorexia is a serious illness with both long- and short-term
increases in mortality. For example, at 6–12 years’ follow-up, the mortality rate is
9.6 times the expected rate (Nielsen, 2001).
The thin ideal of female attractiveness often is thought to be a cultural stress
leading to the development of anorexia. Studies conducted among teenage girls in
Fiji have shown that the introduction of television (with Western programming) in
1995 lead to an increased concern with maintaining a thin body and increased rates
of dieting and purging (Becker et al., 2007). Anorexia in some non-Western cultures
takes a somewhat different form, however. Anorexia patients in Hong Kong do not
have the “fat phobia” we associate with Western anorexia, but rather exhibit a generalized avoidance of eating (Katzman and Lee, 1997). This indicates that even though
anorexia is not limited to Western cultures, the focus on fat (rather than a more generalized obsession with self-control) is shaped by the Western cultural concerns with
obesity, thinness, and weight loss.
Most young women maintain their body weight without starving themselves,
habitually purging, or even dieting. In a 1-year longitudinal study of the eating
and dieting habits of 231 American adolescent girls, medical anthropologist Mimi
Nichter and colleagues (1995) showed that most of the subjects maintained their
weight by watching what they eat and trying to follow a healthful lifestyle rather than
taking more extreme measures. Anthropological studies such as this are important
because clinicians are not as interested in what the healthy population is doing, and
they help to provide a biocultural context for the expression of disease.
Birth , Growth and Aging
All animals go through the processes of birth, growth, and aging. Normal growth
and development are not medical problems per se, but the process of growth is
a sensitive overall indicator of health status (Tanner, 1990). Therefore, studies of
growth and development in children provide useful insights into the nutritional or
environmental health of populations.
Human Childbirth
Nothing should be more natural than giving birth. After all, the survival of the species
depends on it. However, in industrialized societies birth usually occurs in hospitals. Of
the more than 4 million births in the United States in 2000, more than 90% occurred in
hospitals; in 2007, 31.8% of all American births were Cesarean deliveries (Martin et al.,
2010). This rate is not extraordinary among developed countries: it is somewhat higher
than those seen in Europe, but lower than rates in many parts of China and Latin
America (Betrán et al., 2007). In 1900, only 5% of U.S. births occurred in a hospital
(Wertz & Wertz, 1989). At that time, given the high risk of contracting an untreatable
infection, hospitals were seen as potentially dangerous places to give birth.
Human females are not that much larger than chimpanzee females, yet they give
birth to infants whose brains are nearly as large as the brain of an adult chimpanzee
and whose heads are very large compared with the
size of the mother’s pelvis. The easiest evolutionary solution to this problem would be for women
to have evolved larger pelves, but too large a pelvis
would reduce bipedal efficiency. Wenda Trevathan
(1999) points out that the shape as well as the size
of the pelvis is a critical factor in the delivery of a
child. Not only is there a tight fit between the size
of the newborn’s head and the mother’s pelvis, but
the baby’s head and body must rotate or twist as
they pass through the birth canal, which is a process that introduces other dangers (such as the
umbilical cord wrapping around the baby’s neck).
In contrast to humans, birth is easy in the great
apes. Their pelves are substantially larger relative
to neonatal brain size, and the shape of their quadrupedal pelves allows a more direct passage of the
newborn through the birth canal (Figure 15.2).
In traditional cultures, women usually give
birth with assistance from a midwife (almost
always a woman). Trevathan observes that although women vary across cultures in their
reactions to the onset of labor, in almost all
cases, the reaction is emotion-charged and
results in the mother seeking assistance from
others. She hypothesizes that this behavior
is a biocultural adaptation. A human birth is
much more likely to be successful if someone
is present to assist the mother in delivery. Part
of the assistance is in actually supporting the
newborn through multiple contractions as it
passes through the birth canal, but much recent research has shown that the emotional
support of mothers provided by birth assistants
is also of critical importance (Klaus & Kennell,
1997). Such emotional support often is lacking
in contemporary hospital deliveries, although
there has been some effort in recent years to
remedy this situation (Figure 15.3). Recent research has shown that birth for large-brained
Neandertal babies was just as difficult as for
modern humans (Ponce de León et al., 2008). It is interesting to consider the
possibility that Neandertal mothers may have also received support from kin and
others during birth.
Patterns of Human Growth
The study of human growth and development is known as auxology. All animals go
through stages of growth that are under some degree of genetic control. However,
the processes of growth and development can be acutely sensitive to environmental conditions. Thus, patterns of growth that emerge under different environmental
conditions can provide us with clear examples
of biological plasticity (Mascie-Taylor and Bogin,
1995) (Chapter 6).
We chart growth and development using
several different measures including height,
weight, and head circumference. Cognitive
skills, such as those governing the development of language, also appear in a typical
sequence as the child matures. We can also
assess age by looking at dentition or sexual
reproductive capacity. Different parts of the
body mature at different rates (Figure 15.4).
For example, a nearly adult brain size is
achieved very early, whereas physical and
reproductive maturation all come later in
childhood and adolescence.
Stages of Human Growth
In the 1960s, Adolph Schultz (1969) proposed a
model of growth in primates that incorporated
four stages shared by all primates. This model is
presented in Figure 15.5. In general, as life span
increases across primate species, each stage of
growth increases in length as well.
The Prenatal or Gestational Stage The first stage of growth is the prenatal or gestational stage. This begins with conception and ends with the birth of
the newborn. As indicatedin Figure 15.5, gestational length increases across
primates with increasing life span but is not simply a function of larger body
size. Gibbons have a 30-week gestation, compared with the approximately
25-week gestation of baboons, even though gibbons are much smaller. Growth during the prenatal period is extraordinarily rapid. In humans, during the embryonic
stage (first 8 weeks after conception), the fertilized ovum (0.005 mg) increases in
size 275,000 times. During the remainder of the pregnancy (the fetal period), growth
continues at a rate of about 90 times the initial weight (the weight at the end of the
embryonic stage) per week, to reach a normal birth weight of about 3,200 g.
Although protected by the mother both physically and by her immune system,
the developing embryo and fetus are highly susceptible to the effects of some
substances in their environment. Substances that cause birth defects or abnormal
development of the fetus are known as teratogens. The most common human
teratogen is alcohol. Fetal alcohol syndrome (FAS) is a condition seen in children that
results from “excessive” drinking of alcohol by the mother during pregnancy. At this
point, it is not exactly clear what the threshold for excessive drinking is or whether
binge drinking or a prolonged low level of drinking is worse for the fetus (Thackray
and Tifft, 2001). Nonetheless, it is clear that heavy maternal drinking can lead to
the development of characteristic facial abnormalities and behavioral problems in
children. It is estimated that between 0.5 and 5 in 1,000 children have some form
of alcohol-related birth defect. Although they are not teratogens, other substances
in the environment may affect the developing fetus. Pollutants such as lead and
polychlorinated biphenyls may cause low birth weight and other abnormalities.
Infancy, Juvenile Stage, Adolescence, and Adulthood Schultz defined the
three stages of growth following birth—infancy, juvenile stage, and adulthood—
with reference to the appearance of permanent teeth. Infancy lasts from birth until
the appearance of the first permanent tooth. In humans, this tooth usually is the
lower first molar, and it appears at 5–6 years of age. The juvenile stage begins at this
point and lasts until the eruption of the last permanent tooth, the third premolar,
which can occur anywhere between 15 and 25 years of age. Adulthood follows the
appearance of the last permanent tooth.
Tooth eruption patterns provide useful landmarks for looking at stages of growth
across different species of primates, but they do not tell the whole story. Besides
length of stages, there is much variation in the patterns of growth and development
in primate species. Barry Bogin (1999) suggests that the four-stage model of primate
growth is too simple and does not reflect patterns of growth that may be unique to
humans. In particular, he argues that during adolescence, humans have a growth spurt
that reflects a species-specific adaptation.
Bogin places the end of the juvenile period, and the beginning of adolescence,
at the onset of puberty. The word puberty literally refers to the appearance of pubic
hair, but as a marker of growth it refers more comprehensively to the period during
which there is rapid growth and maturation of the body (Tanner, 1990). The age
at which puberty occurs is tremendously variable both within and between populations, and even within an individual, different
parts of the body may mature at different rates
and times. Puberty tends to occur earlier in girls
than boys. In industrialized societies, almost all
children go through puberty between the ages
of 10 and 14 years (Figure 15.6).
During adolescence, maturation of the primary and secondary sexual characteristics continues. In addition, there is an adolescent growth
spurt. According to Bogin (1993, 1999), the
expanding database on primate maturation
patterns indicates that the adolescent growth
spurt—and therefore adolescence—is most pronounced in humans. Why do we need adolescence? The length of the juvenile stage, most of
which occurs after brain size has reached adult
proportions, varies widely among mammal species. There is a cost to a prolonged juvenile stage
because it delays the onset of full sexual maturity and the ability to reproduce. But the juvenile stage is also necessary as a training period
during which younger animals can learn their
adult roles and the social behaviors necessary to survive and reproduce within their own species. The evolutionary costs of delaying
maturation are offset by the benefits of social life. Among mammals, the juvenile
stage is longest in highly social animals, such as wolves and primates. Humans are
the ultimate social animal. Bogin argues that the complex social and cultural life of
humans, mediated by language, requires an extended period of social learning and
development: adolescence.
The Secular Trend in Growth
One of the most striking changes in patterns of growth identified by auxologists is
the secular trend in growth. By using data collected as long ago as the eighteenth century, they demonstrated that in industrialized countries, children have been growing larger and maturing more rapidly with each passing decade, starting in the late
nineteenth century in Europe and North America (Figure 15.7). The secular trend
started in Japan after World War II, and it is just being initiated now in parts of the
developing world. In Europe and North America, since 1900, children at 5 to 7 years
of age averaged an increase in stature of 1 to 2 cm per decade (Tanner, 1990). In
Japan between 1950 and 1970, the increase was 3 cm per decade in 7-year-olds and
5 cm per decade in 12-year-olds. A more recent secular trend in growth has been seen
in South Korea, where surveys of children conducted between 1965 and 2005 show
a continuing increase in both height and weight (Kim et al., 2008). Twenty-year-old
Korean men were 5.3 cm taller and 12.8 kg heavier than their 1965 counterparts;
women were 5.4 cm taller and 4.1 kg heavier. The onset of puberty was clearly earlier
in the 2005 group, since the greatest differences from the 1965 group were seen in
the 10–15-year-old age groups.
The secular trend in growth undoubtedly is a result of better nutrition (more
calories and protein in the diet) and a reduction in the impact of diseases during
infancy and childhood. We find evidence for this over the short term from migration
studies, which have shown that changes in the environment (from a less healthful
to a more healthful environment) can lead to the development of a secular trend
in growth. Migration studies look at a cohort of the children of migrants born and
raised in their new country and compare their growth with either their parents’
growth (if the children have reached adulthood) or that of a cohort of children in
the country from which they immigrated. Migration studies of Mayan refugees from Guatemala to the United States show evidence of a secular trend in growth (Bogin,
1995). Mayan children raised in California and Florida were on average 5.5 cm taller
and 4.7 kg heavier than their counterparts in Guatemala. Although the secular trend
in growth appears to highlight a straightforward relationship between increased stature and industrialization, the stature each individual achieves is the result of the
complex interaction of genetics, economic status, and nutrition.
Menarche and Menopause
Another hallmark of the secular trend in growth is a decrease in the age of
menarche—a girl’s first menstrual period—seen throughout the industrialized
world. From the 1850s until the 1970s, the average age of menarche in European and
North American populations dropped from around 16 to 17 years to 12 to 13 years
(Figure 15.8) (Tanner, 1990; Coleman & Coleman, 2002). A comprehensive study of
U.S. girls (sample size of 17,077) found that the age of menarche was 12.9 years for
white girls and 12.2 years for black girls (Herman-Giddens et al., 1997). This does
not reflect a substantial drop in age of menarche since the 1960s.
In cultures undergoing rapid modernization, changes in the age of menarche
have been measured over short periods of time. Among the Bundi of highland
Papua New Guinea, age of menarche dropped from 18.0 years in the mid-1960s to
15.8 years for urban Bundi girls in the mid-1980s (Worthman, 1999). Over the long
term, the rate of decrease in age of menarche in most of the population was in the
range 0.3 to 0.6 years per decade. For urban Bundi girls, the rate is 1.29 years per
decade, which may be a measure of the rapid pace of modernization in their society.
Menarche marks the beginning of the reproductive life of women, whereas
menopause marks its end. Menopause is the irreversible cessation of fertility that
occurs in all women before the rest of the body shows other signs of advanced aging
(Peccei, 2001a). Returning to Figure 15.5 on page 391, note that of all the primate
species illustrated, only in humans does a significant part of the life span extend
beyond the female reproductive years. In fact, as far as we know, humans are unique
in having menopause (with the exception of a species of pilot whale). Menopause
has occurred in the human species for as long as recorded history (it is mentioned
in the Bible), and there is no reason to doubt that it has characterized older human
females since the dawn of Homo sapiens. Although highly variable, menopause usually
occurs around the age of 50 years.
At first glance, menopause looks to be a
well-defined, programmed life–history stage.
Why does it occur? Jocelyn Peccei (1995)
suggests a combination of factors, including
adaptation, physiological tradeoff, and an artifact of the extended human life span. Some
adaptive models focus on the potential fitness benefits of having older women around
to help their daughters raise their children,
termed the grandmothering hypothesis (Hill &
Hurtado, 1991). Kristen Hawkes (2003) proposes that menopause is the most prominent
aspect of a unique human pattern of longevity and that this pattern has been shaped
largely by the inclusive fitness benefits derived by postmenopausal grandmothers who
contribute to the care of their grandchildren.
There is some empirical support for this idea.
For example, a study of Finnish and Canadian
historical records indicates that women who
had long postreproductive lives had greater
lifetime reproductive success (Lahdenpera
et al., 2004).
Peccei suggests that an alternative to the
grandmothering hypothesis may be more plausible: the mothering hypothesis. She argues that
the postreproductive life span of women allows
them to devote greater resources to the (slowly
maturing) children they already have and that
this factor alone could account for the evolution
of menopause. This hypothesis is supported by
population data from Costa Rica covering maternal lineages dating from the 1500s until the 1900s
(Madrigal and Meléndez-Obando, 2008). These
data showed that the longer a mother lived, the
higher her fitness; however, there was a negative
effect on her daughter’s fitness. Thus there was
support for the mothering hypothesis but not
the grandmothering hypothesis. Clearly, more
research needs to be done in this area. The relationship between maternal longevity and reproductive fitness is complex, and we will need data
from many populations before there is a general
perspective on that relationship in the human
species as a whole.
Aging
Compared with almost all other animal species, humans live a long time, at least as
measured by maximum life span potential (approximately 120 years). But the human
body begins to age, or to undergo senescence, starting at a much younger age. Many
bodily processes actually start to decline in function starting at age 20, although the
decline becomes much steeper starting between the ages of 40 and 50 (Figure 15.9).
The physical and mental changes associated with aging are numerous and well
known, either directly or indirectly, to most of us (Schulz & Salthouse, 1999).
Why do we age? We can answer from both the physiological and the evolutionary standpoints (Figure 15.10). From a physiological perspective, several hypotheses or models of aging have been offered (Nesse & Williams, 1994; Schulz & Salthouse,
1999). Some have focused on DNA, with the idea that over the lifetime, the accumulated damage to DNA, in the form of mutations caused by radiation and other
forces, leads to poor cell function and ultimately cell death. Higher levels of DNA repair enzymes are found in longer-lived species, so there may be some validity to this
hypothesis, although in general the DNA molecule is quite stable. Another model
of aging focuses on the damage that free radicals can do to the tissues of the body
(Finkel and Holbrook, 2000). Free radicals are molecules that contain at least one
unpaired electron. They can link to other molecules in tissues and thereby cause
damage to those tissues. Oxygen-free radicals, which result from the process of oxidation (as the body converts oxygen into energy), are thought to be the main culprit
for causing the bodily changes in aging. Antioxidants, such as vitamins C and E,
may reduce the effects of free radicals, although it is not clear yet whether they slow
the aging process. Further evidence for the free radical theory of aging comes from
diseases in which the production of the body’s own antioxidants is severely limited.
These diseases seem to mimic or accelerate the aging process.
In wild populations, aging is not a major contributor to mortality: Most animals
die of something besides old age, as humans did before the modern age. Thus, aging
per se could not have been an adaptation in the past because it occurred so rarely in
the natural world (Kirkwood, 2002). Two nonadaptive evolutionary models of aging
are the disposable soma hypothesis (Kirkwood and Austad, 2000) and the pleiotropic gene
hypothesis (Williams, 1957; Nesse and Williams, 1994). Both take the position that old
organisms are not as evolutionarily important as young organisms. The disposable
soma hypothesis posits that it is more efficient for an organism to devote resources
to reproduction rather than maintenance of a body. After all, even a body in perfect shape can still be killed by an accident, predator, or disease. Therefore, organisms are better off devoting resources to getting their genes into the next generation
rather than fighting the physiological tide of aging.
The pleiotropic gene hypothesis has a similar logic, although it comes at the problem from a different angle. As you recall (Chapter 4), pleiotropy refers to the fact that
most genes have multiple phenotypic effects. For all organisms, the effects of natural
selection are more pronounced based on the phenotypic effects of the genes during
the earliest rather than later phases of reproductive life. The simple reason for this is
that a much higher proportion of organisms live long enough to reach the early reproductive phase than the proportion who make it until the late reproductive phase.
For example, imagine that a gene for calcium metabolism helps a younger animal
heal more quickly from wounds and thus increase its fertility (Nesse and Williams,
1994). A pleiotropic effect of that same gene in an older animal might be the development of calcium deposits and heart disease; this “aged” effect has little influence on
the lifetime fitness of the animal. Aging itself may be caused by the cumulative actions
of pleiotropic genes that were selected for their phenotypic effects in younger bodies
but have negative effects as the body ages.
Infectious Diseases and Biocultural Evolution
Our bodies provide the living and reproductive environment for a wide variety of
viruses, bacteria, single-celled eukaryotic parasites, and more biologically complex
parasites, such as worms. As we evolve defenses to combat these disease-causing organisms, they in turn are evolving ways to get around our defenses. Understanding
the nature of this “arms race” and the environments in which it is played out may be
critical to developing more effective treatments in the future.
Infectious diseases are those in which a biological agent, or pathogen, parasitizes or infects a host. Human health is affected by a vast array of pathogens. These
pathogens usually are classified taxonomically (such as bacteria or viruses), by their
mode of transmission (such as sexually transmitted, airborne, or waterborne), or by the
organ systems they affect (such as respiratory or brain infections, or “food poisoning” for the digestive tract). Pathogens vary tremendously in their survival strategies.
Some pathogens can survive only when they are in a host, whereas others can persist
for long periods of time outside a host. Some pathogens live exclusively within a
single host species, whereas others can infect multiple species or may even depend
on different species at different points in their life cycle.
Human Behavior and the Spread of Infectious Disease
Human behavior is one of the critical factors in the spread of infectious disease.
Actions we take every day influence our exposure to infectious agents and determine which of them may or may not be able to enter our bodies and cause an illness.
Food preparation practices, sanitary habits, sex practices, whether one spends time in
proximity to large numbers of adults or children—all these can influence a person’s
chances of contracting an infectious disease. Another critical factor that influences
susceptibility to infectious disease is overall nutritional health and well-being. People
weakened by food shortage, starvation, or another disease (such as cancer) are especially vulnerable to infectious illness (Figure 15.11). For example, rates of tuberculosis in Britain started to decline in the nineteenth century before the bacteria that
caused it was identified or effective medical treatment was developed. This decline
was almost certainly due to improvements in nutrition and hygiene (McKeown, 1979).
Just as individual habits play a prominent role in the spread of infectious disease,
so can widespread cultural practices. Sharing a communion cup has been linked to
the spread of bacterial infection, as has the sharing of a water source for ritual washing before prayer in poor Muslim countries (Mascie-Taylor, 1993). Cultural biases
against homosexuality and the open discussion of sexuality gave shape to the entire
AIDS epidemic, from its initial appearance in gay communities to delays by leaders
in acknowledging the disease as a serious public health problem.
Agriculture Agricultural populations are not necessarily more vulnerable to infectious disease than hunter–gatherer populations. However, larger and denser agricultural populations are likely to play host to all the diseases that affect hunter–gatherer
populations and others that can be maintained only in larger populations. This is the
basis of the first epidemiological transition discussed earlier. For example, when a
child is exposed to measles, his or her immune system takes about 2 weeks to develop
effective antibodies to fight the disease. This means that in order to be maintained in
a population, the measles virus needs to find a new host every 2 weeks; in other words,
there must be a pool of twenty-six new children available over the course of a year to
host the measles virus. This is possible in a large agricultural population but almost
impossible in a much smaller hunter–gatherer population (Figure 15.12).
Agriculture and nonagricultural populations also differ in that the former tend
to be sedentary, whereas the latter tend to be nomadic. Large, sedentary agricultural
populations therefore are more susceptible to bacterial and parasitic worm diseases
that are transmitted by contact with human waste products. In addition, many diseases
are carried by water, and agricultural populations are far more dependent on a limited
number of water sources than nonagricultural populations. Finally, agricultural populations often have domestic animals and also play host to a variety of commensal animals, such as rats, all of which are potential carriers of diseases that may affect humans.
Specific agricultural practices may change the environment and encourage the
spread of such infectious diseases as sickle cell and malaria. Slash-and-burn agriculture
leads to more open forests and standing pools of stagnant water. Such pools are an
ideal breeding ground for the mosquitoes that carry the protozoa that cause malaria.
Mobility and Migration The human species is characterized by its mobility. One
price of this mobility has been the transmission of infectious agents from one population to another, leading to uncontrolled outbreaks of disease in the populations
that have never been exposed to the newly introduced diseases. These are referred
to as virgin soil epidemics.
The Black Death in Europe (1348–1350) is one example of just such an outbreak (Figure 15.13). The “Black Death” was bubonic plague, a disease caused by
the bacterium Yersinia pestis. The bacterium is transmitted by the rat flea, which lives
on rats. When the fleas run out of rodent hosts, they move to other mammals, such
as humans. The bacteria can quickly overwhelm the body, causing swollen lymph
nodes (or buboes, hence the name) and in more severe cases lead to infection of the
respiratory system and blood. It can kill very quickly. An outbreak of bubonic plague
was recorded in China in the 1330s, and by the late 1340s it had reached Europe.
In a single Italian city, Florence, a contemporary report placed the number dying between March and October 1348 at 96,000. By the end of the epidemic, one-third
of Europeans (25–40 million) had been killed, and the economic and cultural life of
Europe was forever changed.
Similar devastation awaited the native peoples of the New World after
1492 with the arrival of European explorers and colonists. Measles, smallpox,
influenza, whooping cough, and sexually transmitted diseases exacted a huge toll
on native populations throughout North and South America, the Island Pacific,
and Australia. Some populations were completely wiped out, and others had
such severe and rapid population depletion that their cultures were destroyed.
In North America, for example, many communities of native peoples lost up
to 90% of their population through the introduction of European diseases
( Pritzker, 2000). Infectious diseases often reached native communities before
the explorers or colonizers did, giving the impression that North America was an
open and pristine land waiting to be filled.
Infectious Disease and the Evolutionary Arms Race
As a species, we fight infectious diseases in many ways. However, no matter what we
do, parasites and pathogens continuously evolve to overcome our defenses. Over
the last 50 years, it appeared that medical science was gaining the upper hand on
infectious disease, at least in developed countries. However, despite real advances,
infectious diseases such as the virus that causes AIDS and antibiotic-resistant bacteria
remind us that this primeval struggle will continue.
The Immune System One of the most extraordinary biological systems that has
ever evolved is the vertebrate immune system, the main line of defense in the fight
against infectious disease. At its heart is the ability to distinguish self from nonself. The
immune system identifies foreign substances, or antigens, in the body and synthesizes
antibodies, which comprise a class of proteins known as immunoglobulins, which are
specifically designed to bind to and destroy specific antigens (Figure 15.14).
The immune system is a complex mechanism that has evolved to deal
with the countless number of potential antigens in the environment. An
example of what happens when just one of the components of the immune
system is not functioning occurs in AIDS. The human immunodeficiency virus
(HIV) that causes AIDS attacks the helper T cells. As mentioned earlier,
the helper T cells respond to antigens by inducing the B lymphocytes to
produce antibodies, leading to the production of phagocytes; when their
function is compromised, the function of the entire immune system is also
compromised. This leaves a person with HIV infection vulnerable to a host
of opportunistic infections, a condition that characterizes the development
of full-blown AIDS.
Cultural and Behavioral Interventions Although the immune
system does a remarkable job fighting infectious disease, it is obviously
not always enough. Even before the basis of infectious diseases was
understood, humans took steps to limit their transmission. Throughout
the Old World, people with leprosy were shunned and forced to live apart
from the bulk of the population. This isolation amounted to quarantine,
in recognition of the contagious nature of their condition.
One of the most effective biocultural measures developed to fight infectious diseases is vaccination. The elimination of smallpox as a scourge of
humanity is one of the great triumphs of widespread vaccination. Smallpox
is a viral illness that originated in Africa some 12,000 years ago and subsequently spread throughout the Old World (Barquet & Domingo, 1997).
It was a disfiguring illness, causing pustulant lesions on the skin, and it
was often fatal. Smallpox killed millions of people upon its introduction
to the New World; in the Old World, smallpox epidemics periodically decimated entire populations. In 180 CE, a smallpox epidemic killed between 3.5 and 7 million
people in the Roman Empire, precipitating the first period of its decline. Crude vaccination practices against smallpox were developed hundreds of years ago, but these often carried a significant risk of developing the disease. During the twentieth century,
modern vaccination methods worked to virtually eliminate this disease (Figure 15.15).
The most recently developed forms of intervention against infectious disease are
drug based. The long-term success of these drugs will depend on the inability of the
infectious agents to evolve resistance to their effects. Overuse of anti-infectious drugs
may actually hasten the evolution of resistant forms by intensifying the selection
pressures on pathogens.
Evolutionary Adaptations The immune system is the supreme evolutionary
adaptation in the fight against infectious disease. However, specific adaptations to
disease that do not involve the immune system are also quite common ( Jackson,
2000). For example, a class of enzymes known as lysozymes attacks the cell wall
structure of some bacteria. Lysozymes are found in high concentrations in the tear
ducts, salivary glands, and other sites of bacterial invasion.
The sickle cell allele has spread in some populations because it functions as an
adaptation against malaria. Another adaptation to malaria is the Duffy blood group
(see Chapter 6). In Duffy-positive individuals, the proteins Fya and Fyb are found on
the surface of red blood cells. These proteins facilitate entry of the malaria-causing
protozoan Plasmodium vivax. Duffy-negative individuals do not have Fya and Fyb on
the surface of their red blood cells, so people with this phenotype are resistant to
vivax malaria. Many Duffy-negative people are found in parts of Africa where malaria
is common; others, who live elsewhere, have African ancestry
Diet and Disease
It seems that there are always conflicting reports on what particular parts of our diet
are good or bad for us. Carbohydrates are good one year and bad the next. Fats go in
and out of fashion. From a biocultural anthropological perspective, American attitudes toward diet and health at the turn of the twenty-first century provide a rich source of
material for analysis. However, despite all the confusion about diet, we all have the same
basic nutritional needs. We need energy (measured in calories or kilojoules) for body
maintenance, growth, and metabolism. Carbohydrates, fat, and proteins are all sources
of energy. We especially need protein for tissue growth and repair. In addition to energy,
fat provides us with essential fatty acids important for building and supporting nerve
tissue. We need vitamins, which are basically organic molecules that our bodies cannot
synthesize but that we need in small quantities for a variety of metabolic processes. We
also need a certain quantity of inorganic elements, such as iron and zinc. For example,
with insufficient iron, the ability of red blood cells to transport oxygen is compromised,
leading to anemia. Finally, we all need water to survive.
The Paleolithic Diet
For most of human history, people lived in small groups and subsisted on wild foods
that they could collect by hunting or gathering. Obviously, diets varied in different
areas: Sub-Saharan Africans were not eating the same thing as Native Americans
on the northwest Pacific coast. Nonetheless, S. Boyd Eaton and Melvin Konner
(Eaton & Konner, 1985; Eaton et al., 1999) argue that we can reconstruct an average
Paleolithic diet from a wide range of information derived from paleoanthropology,
epidemiology, and nutritional studies. A comparison of the average Paleolithic and
contemporary diets is presented in Table 15.1 (Eaton et al., 1999).
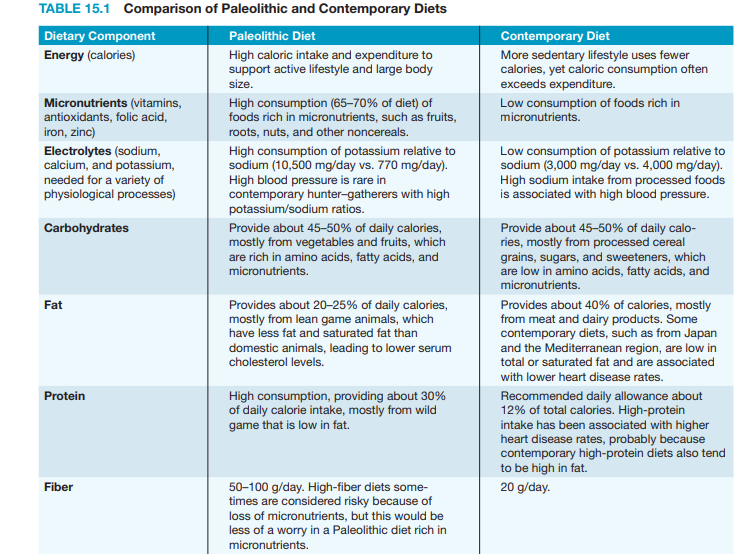
The contemporary diet is not simply a more abundant version of the hunter–
gatherer diet. It differs fundamentally in both composition and quality. Compared
with contemporary diets, the hunter–gatherer diet can be characterized as being
high in micronutrients, protein, fiber, and potassium and low in fat and sodium.
Total caloric and carbohydrate intake is about the same in both diets, but hunter–
gatherers typically were more active than contemporary peoples and thus needed
more calories, and their carbohydrates came from fruits and vegetables rather than
processed cereals and refined sugars.
The comparison between hunter–gatherer and contemporary diets indicates that
increasing numbers of people are living in nutritional environments for which their
bodies are not necessarily well adapted. With few exceptions (such as the evolution
of lactose tolerance) there has not been enough time, or strong enough selection
pressures, for us to develop adaptations to this new nutritional environment. Indeed,
because most of the negative health aspects of contemporary diets (obesity, diabetes,
cancer) become critical only later in life, it is likely that health problems associated
with the mismatch between our bodies and our nutritional environment will be with
us for some time.
Agriculture and Nutritional Deficiency
Agriculture allowed the establishment of large population centers, which in turn
led to the development of large-scale, stratified civilizations with role specialization.
Agriculture also produced an essential paradox: From a nutritional standpoint,
most agricultural people led lives that were inferior to the lives of hunter–gatherers.
Agricultural peoples often suffered from nutritional stress as dependence on a few
crops made their large populations vulnerable to both chronic nutritional shortages
and occasional famines. The “success” of agricultural peoples relative to hunter–
gatherers came about not because agriculturalists lived longer or better lives but because there were more of them.
With their dependence on a single staple cereal food, agricultural populations
throughout the world have been plagued by diseases associated with specific nutritional deficiencies. As in the Illinois Valley, many populations of the New World were
dependent on maize as a staple food crop. Dependence on maize is associated with
the development of pellagra, a disease caused by a deficiency of the B vitamin niacin
in the diet. Pellagra causes a distinctive rash, diarrhea, and mental disturbances, including dementia. Ground corn is low in niacin and in the amino acid tryptophan,
which the body can use to synthesize niacin. Even into the twentieth century, poor
sharecroppers in the southern United States and poor farmers in southern Europe,
both groups that consumed large quantities of cornmeal in their diets, were commonly afflicted with pellagra. Some maize-dependent groups in Central and South
America were not so strongly affected by pellagra because they processed the corn
with an alkali (lye, lime, ash) that released niacin from the hull of the corn.
In Asia, rice has been the staple food crop for at least the last 6,000 years. In
China, a disease we now call beriberi was first described in 2,697 BCE. Although it was
not recognized at that time, beriberi is caused by a deficiency in vitamin B1 or thiamine. Beriberi is characterized by fatigue, drowsiness, and nausea, leading to a variety
of more serious complications related to problems with the nervous system (especially
tingling, burning, and numbness in the extremities) and ultimately heart failure. Rice
is not lacking in vitamins; however, white rice, which has been polished and milled to
remove the hull, has been stripped of most of its vitamin content, including thiamine.
Agriculture and Abundance: Thrifty and Nonthrifty Genotypes
The advent of agriculture ushered in a long era of nutritional deficiency for most
people. However, the recent agricultural period, as exemplified in the developed
nations of the early twenty-first century, is one of nutritional excess, especially in terms of the consumption of fat and carbohydrates of little nutritional value
other than calories. The amount and variety of foods available to people in
contemporary societies are unparalleled in human history.
In 1962, geneticist James Neel introduced the idea of a thrifty genotype, a
genotype that is very efficient at storing food in the body in the form of fat,
after observing that many non-Western populations that had recently adopted
a Western or modern diet were much more likely than Western populations
to have high rates of obesity, diabetes (especially Type 2 or non–insulindependent diabetes), and all the health problems associated with those conditions (see also Neel, 1982). Populations such as the Pima-Papago Indians in
the southwest United States have diabetes rates of about 50%, and elevated
rates of diabetes have been observed in Pacific Island-, Asian-, and Africanderived populations with largely Western diets (Figure 15.16).
According to Neel, hunter–gatherers needed a thrifty genotype to adapt
to their nonabundant nutritional environments; in contrast, the thrifty
genotype had been selected against in the supposedly abundant European
environment through the negative consequences of diabetes and obesity. The
history of agriculture and nutritional availability in Europe makes the evolution of a nonthrifty genotype unlikely (Allen & Cheer, 1996); Europe was no
more nutritionally favored than other agricultural or hunter–gatherer populations.
However, the notion of a thrifty genotype retains validity. At its heart is the idea that
we are adapted to a lifestyle and nutritional environment far different from those we
find in contemporary populations.
We have seen how growth patterns, infectious disease exposure, nutritional
status, and a host of other health-related issues are fundamentally changed by the
adoption of new cultural practices and technologies. Conditions such as rickets and
diabetes have been called diseases of civilization because they seem to be a direct
result of the development of the industrialized urban landscape and food production. But this label is misleading. Rickets could also be called a disease of migration
and maladaptation to a specific environment. Diabetes could be characterized as a
disease of nutritional abundance, which was certainly not a characteristic of civilization for most of human history.
Biomedical anthropology is interested in understanding the patterns of human
variation, adaptation, and evolution as they relate to health issues. This entails an
investigation of the relationship between our biologies and the environments we live
in. Understanding environmental transitions helps us understand not only the development of disease but also the mechanisms of adaptation that have evolved over
thousands of years of evolution. Change is the norm in the modern world. In the
future, we should expect human health to be affected by these changes. By their
training and interests, biological anthropologists will be in an ideal position to make
an important contribution to understanding the dynamic biocultural factors that
influence human health and illness.
Forensic Anthropology Life, Death and Skeleton
The field of forensic anthropology has achieved recent popularity due in part to
television shows such as CSI and Bones. But like most popularizations, the fantasy is
more glamorous than the reality. Each state has medical examiners or coroners who
are legally responsible for signing death certificates and determining the cause and
manner of death of people not in the care of a doctor. They also have the authority
to consult other experts in their investigations, including forensic anthropologists.
A forensic anthropologist is often consulted in cases in which soft-tissue remains are
absent or badly decomposed.
Forensic anthropologists work with clues about growth, health, disease and
adaptation that are visible in each of our skeletons. These scientists are specialists in human osteology who use the theory and method of
biological anthropology to answer questions about how
recent humans lived and died. They study skeletal remains from crime scenes, war zones, and mass disasters
within the very recent past to reveal the life history of the
individual, to identify that individual, and to understand
something about the context in which death occurred.
They use osteological identification and archaeological field methods to retrieve remains and to develop a
profile of the age, sex, and other biological attributes of
an individual. Because the way the skeleton of a human
or any other animal looks is dictated by its function in life
and its evolutionary history, forensic anthropologists can
reconstruct the probable age, sex, and ancestry of an individual from his or her skeletal remains. They can observe
the influence of certain kinds of diseases on the skeleton,
and they can assess some aspects of what happened to an
individual just before, around the time of, and after his
or her death. Unlike pathologists and medical examiners,
forensic anthropologists bring an anthropological perspective and a hard-tissue
focus to investigations of skeletal remains.
Field Recovery and Laboratory Processing
Forensic investigations rely on good contextual information. Things like body position, relationship to nearby items such as bullets or grave goods, and structures near
the individual require precise and thorough documentation in the field. Without
such documentation, we would not know if the bullet recovered at the scene was
10 feet from the individual, or within the victim’s chest cavity. These associations are
crucial for inferring the meaning of a burial and the circumstances surrounding the
death of an individual. So to ensure full recovery and good contextual information,
we rely on archaeological techniques to find, document, and remove remains from
the site. When the site is identified it is cordoned off and surveyed for additional
remains. Such surveys commonly include an individual or team of investigators walking a systematic path searching for remains, associated items, or evidence of burial
(Figure 15.17).
In the field, any surface discoveries are mapped and photographed. A permanent datum point for the site is established that represents a fixed position from
which everything is measured so that the precise “find spot” of each object can be
relocated in the future. If the remains are buried, the anthropologist will excavate
using archaeological techniques. The excavator begins by skimming off shallow layers of dirt using a hand trowel. Objects are revealed in place and their coordinates,
including their depth, are recorded relative to the grid system (Figure 15.18), and
photographs are taken. The dirt is sieved through fine mesh to ensure that even
the smallest pieces of bone are recovered (Figure 15.19). Soil samples may be saved
to assist in the identification of insects and plants. In the field, the anthropologist
makes a preliminary determination of whether the remains are human or nonhuman (they could be those of a dog or deer, for instance) and, based on the bones,
whether more than one individual is present. Once exposed and mapped, individual
bones are tagged, bagged, and removed to the laboratory.
In the lab more detailed curation and examination can begin. A strict
chain of custody is established to ensure that the remains cannot be tampered with,
in case they should become evidence in a court of law. Detailed notes are taken to
demonstrate that the remains in question are those from the scene, that they have
not been contaminated or modified since their removal from the scene, and who has
had access to them.
After the remains are cataloged, they are cleaned of any adhering soft tissue and dirt, and then laid out in anatomical position, the
way they would have looked in the skeleton in life (Figure 15.20).
An inventory is made of each bone present and its condition. Most
adult humans have 206 bones, many of which are extremely small
(see Appendix A). Because most bones develop as several bony centers that fuse together only later in life, fetuses and children contain
many more bones than do adults. Often the scientist works with no
more than a few bone fragments.
The Biological Profile
Once the initial inventory has been completed, the scientist sets
about evaluating the clues that the skeleton reveals about the
life and death of the individual. The first step in this process is
constructing the biological profile of the individual— including
determining age, sex, height, and disease status.
Age at Death
As the human body develops, from fetus to old age, dramatic changes occur throughout the skeleton. Scientists use the more systematic of these changes to estimate the
age at death of an individual. However, whenever scientists determine age, they
always report it as a range (such as 35–45 years) rather than as a single definitive
number. This range reflects the variation in growth and aging seen in individuals
and across human populations and denotes the person’s biological rather than
chronological age (age in years). The goal is that the range also encompasses the
person’s actual age at the time of their death.
Because the skeleton grows rapidly during childhood, assessing the age of a
subadult younger than about 18 years of age is easier and often more precise than
estimating the age of an adult skeleton. Virtually all skeletal systems except the small bones of the ear (the ear ossicles) change from newborn to adult. For example,
in small children the degree of closure of the cranial bones (covering the fontanelles, or “soft spots” of the skull) changes with age, as does the development of the
temporal bone, the size and shape of the wrist bones, and virtually every other bone
(Figure 15.21). However, dental eruption and the growth of long bones are the most
frequently used means of assessing subadult age.
Humans have two sets of teeth of different sizes that erupt at fairly predictable
intervals. Which teeth are present can help distinguish between children of different
ages and between older subadults and adults of the same size (Figure 15.22). For
more precise ages, the relative development of the tooth roots can also be used.
However, once most of the adult teeth have erupted, by about the age of 12 years in
humans, the teeth are no longer as good a guide to predicting age.
In these older children, growth of the limb bones can also be used to assess
age. The long bones of the arms and legs have characteristic bony growths at
each end—the epiphyses—which are present as separate bones while the person
is still growing rapidly (Figure 15.23). Most epiphyses are not present at birth—
which helps to separate fetuses from newborns—but appear during infancy and
childhood. The lengths and proportions of bones change in predictable ways as
children grow and are especially good indicators for assessing fetal age ( Sherwood
et al., 2000). In older children, the epiphyses start to fuse to the shafts of the
limb bones around the age of 10 in some bones, and fusion of most epiphyses is
completed in the late teenage years. However, the process of fusion may occur as
late as the early twenties in a few bones (such as the clavicle). Depending upon
which bones and which parts of those bones are fused, a reasonably good estimate
of subadult age can be made.
In adults, age is harder to determine because growth is essentially complete.
Some of the last epiphyses to fuse, such as the clavicle and top of the ilium, can be
used to estimate age in young adults in their early twenties. But estimating the age
of the older adult skeleton relies mostly on degeneration of parts of the skeleton.
For example, the pubic symphysis and auricular surface of the innominate, and
the end of the fourth rib near the sternum all show predictable changes with age
(Todd, 1920, 1921; McKern & Stewart, 1957; Iscan et al., 1984; Lovejoy et al., 1985).
Examination of as many of these bones as possible helps to increase age accuracy(Bedford et al., 1993). The pubic symphysis is a particularly useful indicator of
adult age, and age standards have been developed separately for males and females (Gilbert & McKern, 1973; Katz & Suchey, 1986; Brooks & Suchey, 1990).
The standards show how the symphysis develops from cleanly furrowed to more
granular and degenerated over time (Figure 15.24). These changes tend to occur
more quickly in females than in males due to the trauma the symphysis experiences during childbirth.
The degree of obliteration of cranial sutures (the junction of the different
skull bones) can also give a relative sense of age—obliteration tends to occur in
older individuals (Lovejoy & Meindl, 1985). The antero–lateral sutures of the skull
are the best for these purposes. However, the correlation between degree of obliteration and age is not very close, and the age ranges that can be estimated are wide.
Sex
If certain parts of the skeleton are preserved, identifying biological sex is easier
than estimating age at death, at least for adults. The two parts of the skeleton that
most readily reveal sex are the pelvis and the skull, and sex characteristics are
more prominent in an adult skeleton than in a child. Humans are moderately sexually dimorphic, with males being larger on average than females (see Table 10.2
on page 258). But their ranges of variation overlap so that size alone cannot separate male and female humans.
The best skeletal indicator of sex is the pelvis. Because of selective pressures
for bipedality and childbirth, human females have evolved pelves that provide a relatively large birth canal (see Chapter 10). This affects the shape of the innominate
and sacrum in females; the pubis is longer, the sacrum is broader and shorter, and
the sciatic notch of the ilium is broader in females than in males (Figure 15.25). The
method is highly accurate (Rogers & Saunders, 1993) because the pelvis reflects directly
the different selective pressures that act on male versus female bipeds. Thus the pelvis
is considered a primary indicator of the sex of the individual. And because the femur
has to angle inward from this wider female pelvis to the knee (to keep the biped’s foot
under its center of gravity; see Chapter 10), the size and shape of the femur also differentiate males and females fairly well (Porter, 1995).
The skull is also a useful indicator of sex, at least in adults. Around puberty, circulating hormones lead to so-called secondary sex characters such as distribution of
body and facial hair. During this time male and female skulls also diverge in shape.
Male skulls are more robust on average than female skulls of the same population.
However, these differences are relative and population dependent; some human
populations are more gracile than others. The mastoid process of the temporal bone
and the muscle markings of the occipital bone tend to be larger in males than in
females, and the chin is squarer in males than in females (Figure 15.25). The browridge is less robust and the orbital rim is sharper in females than in males, and the
female frontal (forehead) is more vertical. These differences form a continuum and
provide successful sex estimates in perhaps 80 to 85% of cases when the population
is known.
Ancestry
Knowing the ancestry of an individual skeleton is important for improving the
accuracy of sex, age, and stature estimates. There is no biological reality to the idea
of fixed biological races in humans (see Chapter 6), but we have learned that the
geographic conditions in which our ancestors evolved influence the anatomy of
their descendants. The term ancestry takes into account the place of geographic
origin, which corresponds to biological realities in ways that the term race does not.
Nonetheless, because of the way in which variation is distributed in humans (there
is more variation within than between groups, and many variation clines run in
directions independent of one another) assessing ancestry from the skeleton is less
accurate than assessing age or sex, and the process is also highly dependent on the
comparativegroups used.
Forensic anthropologists base ancestry assessments on comparisons with
skeletal populations of known ancestry. An isolated skull can be measured and
compared using multivariate statistics with the University of Tennessee Forensic
Data Bank of measurements from crania of known ancestry. This process provides
a likely assignment of ancestry and a range of possible error. However, human
variation is such that many people exist in every population whose skulls do not
match well with most other skulls of similar geographic origin. Nonetheless, the
ability to even partially assign ancestry can be useful in several forensic contexts.
Missing person reports often provide an identification of ancestry, and although
this is not based directly on the skeleton, a skeletal determination of ancestry may
suggest a match that could be confirmed by other more time–consuming means
such as dental record comparisons or DNA analysis (see Innovations: Ancestry and
Identity Genetics on pages 410–411).
Innovations On Ancestroy and Genetics
Genetic studies have long
been used for
tracing the histories of
populations (Chapters
6 and 13). As geneticists have discovered
an increasing variety
of markers that are associated with specific
geographical regions
and populations, the
ability to trace individual genetic histories
has increased greatly,
and the ability to make
direct matches to DNA from a crime scene has become an
important forensic technique. In addition, the development
of technologies allowing direct sequencing of DNA regions
quickly and relatively inexpensively means that anyone
can obtain a genetic profile in a matter of a few weeks.
There are two basic approaches to determining personalized genetic histories (PGHs) (Shriver and Kittles,
2004). The first one is the lineage-based approach. These
are based on the maternally inherited mtDNA genomes
and the paternally inherited Y chromosome DNA. The
lineage-based approach has been very useful for population studies, and allows individuals to trace their ultimate
maternal and paternal origins. For example, African American individuals can find out what part of Africa their founding American ancestors may have come from (http://www
.african-ancestry.com). These are the same techniques
that have been used to consider the dispersal and migration of ancient and recent peoples. For example, in a survey of more than 2,000 men from Asia using more than 32 genetic markers, Tatiana Zerjal and her colleagues (2003)
found a Y chromosome lineage that exhibited an unusual
pattern thought to represent the expansion of the Mongol
Empire. They called this haplotype the star cluster (reflecting the emergence of these similar variants from a common source). The star cluster lineage is found in sixteen
different populations, distributed across Asia from the Pacific Ocean to the Caspian Sea. The MRCA (most recent
common ancestor) for this cluster was dated to about 1,000
years ago, and the distribution of populations in which the
lineage is found corresponds roughly to the maximum extent of the Mongol Empire. The Empire reached its peak
under Genghis Khan (c. 1162–1227) and Khan and his close
male relatives are said to have fathered many children
(thousands, according to some historical sources).
One additional population outside the Mongol Empire
also has a high frequency of the star cluster: the Hazaras
of Pakistan (and Afghanistan), many of whom through oral
tradition consider themselves to be direct male-line descendants of Genghis Khan. The star cluster is absent from other
Pakistani populations. The distribution of the star cluster
could have resulted from the migration of a group of Mongols
carrying the haplotype or may even reflect the Y chromosome carried specifically by Genghis Khan and his relatives.
From the perspective of determining an individual’s
PGH, however, the lineage-based approach is limited because it traces only the origins of a very small portion of an
individual’s genome and does not reflect the vast bulk of a
person’s genetic history. In contrast to the lineage-based
approach, autosomal marker-based tests use information
from throughout the genome. Ancestry informative markers
(AIMs) are alleles on the autosomal chromosomes that show
substantial variation among different populations. The more
AIMs that are examined in an individual, the more complete
the picture of that individual’s biogeographical ancestry can be obtained (Shriver and Kittles,
2004). Combining the information from all of these AIMs requires some major statistical
analysis, which has to take into
account the expression of each
marker and its population associations. There will be some
statistical noise in the system
due to factors such as the overlapping population distribution
of the markers and instances of
convergent evolution. In addition, even when a hundred markers are used, the tests sample only a small portion of your genome that is the product
of the combined efforts of thousands of ancestors. The biogeographical ancestry of a person, expressed in terms of percentage affiliations with different populations, is a statistical
statement, not a direct rendering of a person’s ancestry. And
both AIMs and lineage-based tests are limited by the comparative samples that form the basis of our knowledge about
the distribution of DNA markers. Thus, if you submit a cheek
swab to several different companies with different comparative databases, you will get somewhat different ancestry results. Nonetheless, they provide us with an intriguing
snapshot of the geographic origins of a person’s ancestors.
Several commercial companies are now in the ancestry
genetics business. We contacted one of these companies,
DNAPrint Genomics (http://www.-AncestryByDNA.com),
and obtained the biogeographical ancestry of two of the
authors of this text, Craig Stanford (CS) and John S. Allen
(JSA). The genetic testing product used is called AncestryByDNA 2.5, which provides a breakdown of an individual’s
PGH in terms of affiliations with four major geographical
groups: European, Native (aka Indigenous) American, SubSaharan African, and East Asian. It combines information
derived from about 175 AIMs.
John Allen’s results were: 46% European, 46% East
Asian, 8% Native American, and 0% Sub-Saharan African.
These results squared quite well with his known family history: His mother was Japanese and his father was an American of English and Scandinavian descent. The 8% Native
American could have come from one or more ancestors on
his father’s side (some of whom arrived in the United States
in the early colonial period). However, the 95% confidence intervals of the test indicate that
for people of predominantly
European ancestry, a threshold
of 10% Native American needs
to be reached before the result
is statistically significant. For
people of predominantly East
Asian descent, the threshold is 12.5%. Therefore, in the
absence of a family history of
Native American ancestry, it
is best to consider the 8% as
statistical noise.
Craig Stanford’s results were: 82% European, 14%
Native American, 4% Sub-Saharan African, and 0% East
Asian. The Native American result, which easily exceeds
the statistical threshold, was a real surprise because CS
has no family history of Native American ancestry. Following this result, his father was tested and was found to have
91% European and 9% Sub-Saharan African ancestry. Thus,
all of CS’s Native American ancestry was derived from his
mother’s side. Although she was not tested, it is reasonable
to conclude that her Native American percentage would
be greater than 25%—the equivalent of a grandparent, although this does not have to represent the contribution of
a single individual. CS found this result to be somewhat
ironic because earlier generations of women on his mother’s
side of the family had been proud members of the Daughters of the American Revolution, a lineage-based organization that was once (but is no longer) racially exclusionary.
Stanford also requested a more detailed European ancestry
genetic test (EuroDNA 1.0). Along with European ancestry,
the tests showed 12% Middle Eastern ancestry. One of his
paternal grandparents was from Italy, and the ancestry of
southern Europeans often reflects population movements
around the Mediterranean Sea, including Middle Eastern
markers. In addition, there has been a long history of some
gene flow from Sub-Saharan Africa into North Africa and
the Middle East, which could explain his father’s statistically significant Sub-Saharan African ancestry.
Ancestry genetics opens windows to the past, but in
some cases, it raises more questions than answers about
where you came from. This is not surprising because we
know that the pattern of genetic variation across all humans is a complex one that does not partition well into regional or “racial” groups, that most of the genetic variation
within humans exists within rather than between groups,
and that different characteristics often follow cross-cutting
clines. We can attest, however, that for anyone interested
in their own biological ancestry, getting a personalized
genetic history can be an exciting experience. Incidentally,
humans are not the only species whose biological past
can be explored: Genetic ancestry testing for dogs is also
becoming available (http://www.whatsmydog.com) and
paternity testing is available for both cats and dogs (http://
www.catdna.org; http://www.akc.org/dna).
Height and Weight
Physical stature reflects the length of the bones that contribute to a person’s height.
Different body shapes have evolved in response to different climatic pressures (see
the ecological rules described by Bergmann and Allen in Chapter 6). Thus, height
and weight estimates will be more accurate if the population of the individual is
known. These differences in proportions relate to differences in bone lengths; as
a result, some populations will tend to have more of their stature explained by leg
length, and some by torso length, for example.
The best estimates of stature from the skeleton are based on summing the
heights of all the bones in the skeleton that contribute to overall height including the cranium, vertebral column, limb, and foot bones (Fully, 1956). This socalled Fully method is fairly accurate, but requires a complete skeleton, a rarity in
archaeological or forensic contexts. Biological anthropologists have developed formulae, which vary by population, for estimating stature based on the length of a
single or several long bones, so that the femur, tibia, or even humerus can be used to
predict stature. These methods use the relationship between the limb bones and the
height in skeletal remains of individuals of known stature to predict stature for an
unknown individual (e.g., Trotter, 1970). For even more incomplete remains, there
are formulae for estimating total length of a long bone from a fragment of that bone
(e.g., Steele & McKern, 1969). The estimated length can then be used to estimate
height—although the error margin increases with each estimate. Like age, stature is
estimated as a range (for example, 5¿10¿¿ to 6¿0¿¿) that hopefully captures the person’s
true height at the time of death.
As you might expect, weight is more difficult to predict since it can vary quite
a lot over an individual’s life time. Nonetheless, formulae exist for predicting the
approximate weight of an individual from his or her weight-bearing joints, such as
the head of the femur. Using the entire skeleton, scientists can estimate body weight
based on formulae that relate height and body breadth to weight in populations of a
different build (Ruff, 2000). Some of these estimates also form the basis for inferring
body size and weight in earlier hominins (see Chapters 11 and 12).
Premortem Injury and Disease
Injuries and sickness suffered during life are also an important part of the biological
profile, and critical for understanding an individual’s life and, perhaps, identity. Not
all diseases or injuries leave marks on bone. However, we can distinguish the ones that
do as having occurred while the person was alive because the bones show evidence of
healing and remodeling (Figure 15.26[b]). Arthritis and infections of bone show up
clearly in skeletal remains. Old healed injuries, such as broken limbs and even gunshot wounds that a person survived for several weeks, also leave their mark.
Premortem fractures can be key evidence of lifeways (Figure 15.26[a]). In forensic
anthropology, multiple healed fractures—especially of the ribs and those typical of defensive wounds—can establish a series of episodes of violence, as is often the case in
child abuse (Walker et al., 1997). Old injuries can also be matched to premortem Xrays
taken when a victim sought medical attention, and thereby help to establish identity.
In addition to injury and disease, lifestyle may leave an indelible mark on the
skeleton; an athlete who uses one side of the body for intense activity (such as a baseball pitcher or tennis player) will have a more robustly developed arm on that side—
especially if he or she began the activity during childhood and continued through
adulthood. Other repetitive activities also cause bone deposition to differ systematically between individuals. Injury, disease, and lifestyle all leave clues on the skeleton
that tell the story of an individual’s life. The skeleton may also be modified by events
that occur well after or around the time of death. These changes can be critically important for understanding the context of death, but to be of use the scientist must be
able to distinguish premortem bone changes from those that happened later in time.
Perimortem and Postmortem Trauma
For anthropologists, perimortem trauma is the physical evidence of activity that happened slightly before, during, or slightly after the time of death. We can differentiate it from premortem injury because in perimortem trauma no healing is evident.
We can distinguish perimortem from postmortem trauma that happened well after
death, because bones retain a large percentage of their organic component during
the perimortem interval. As a result, they are more pliable and break differently
than those that are well dried out after death; think of the difference between how a
small branch that has just been plucked from a tree bends when you try to break it,
whereas a long-dead, dried-out stick is brittle and snaps in two.
Distinguishing perimortem trauma is one of the most routine tasks a forensic
anthropologist undertakes. This type of evidence helps investigators understand what
happened right around the time of death. This information also helps the medical examiner or coroner determine the cause and manner of death (whether homicide, suicide, or accident) and may help to establish intent in murder cases. For example, the
presence of telltale fractures of the hyoid, a small bone in the neck, suggests strangulation. Perimortem trauma may also indicate a perpetrator’s intent to hide or dispose of
a body, implying that death was not accidental. Circular saws and reciprocating saws
are often used to dismember bodies after a murder. These tools leave different marks
on bone and sometimes leave traces of metal fragments embedded in bone. Experts
can identify types of blades used and can indicate whether they are in the same class
of tools as those owned by a suspect. Being able to show, based on anatomical knowledge, that a body was fleshed when dismembered rather than skeletonized has serious
implications for inferring a crime or interpreting mortuary ritual in past societies.
Definitively postmortem events are not related to establishing cause and manner of death and are often of greater interest to bioarchaeologists than they are to
forensic anthropologists. Analyses of postmortem events can be critical for establishing how bones arrived at a site: Were they deliberately placed in a burial cave, or
did the individual unceremoniously fall through a chasm in the rock? Despite their
greater importance for bioarchaeologists, postmortem events may rule out a crime
if they suggest that marks on bone are made by natural causes, rather than knives,
guns, or chainsaws, or if they show that the skeleton is of ancient rather than forensic
interest. For instance, Willey and Leach (2003) cite a case in which forensic anthropologists sought to identify a human skull found in a suburban home. The skull was
discolored in a variety of ways that most closely resembled the way in which skulls are
sometimes treated when collected as trophies of war. As it turned out, the skull in
question was a “souvenir” brought home from the Vietnam War by a man who had
since moved away, leaving the skull in his garage.
Identification and Forensic Anthropology
Ideally, forensic anthropologists are trying to establish the identity of a victim. To do
this they first develop the biological profile to narrow the field of focus of potential
identities, and they define the time frame of the event. Once they have several possibilities, they can compare a number of different antemortem records to try to establish
an identification. The most common are dental records, surgical implants, and the
matching of antemortem and postmortem Xrays. But DNA can sometimes be used.
DNA analysis has given forensic scientists a powerful new tool for identifying
victims and establishing the presence of an alleged perpetrator at a crime scene.
However, there are also limitations to each of these uses. DNA testing can use tiny
samples of hair, skin, blood, other body fluids, and even bone. However, the older
the bone sample and the more hot and humid the environment in which it was buried, the less likely it is that DNA can be extracted from bone. Forensic scientists use
a variety of tests, including DNA profiling (examining gene sequences that only kin
would be likely to share), DNA typing (isolating particular segments of the genetic
sequence for analysis), and DNA fingerprinting (the original DNA test, in which the
same segments of DNA are lined up to examine the degree of similarity between
samples from two people) (Nafte, 2000).
When skeletal material has been fragmented during a disaster (as in the World
Trade Center crime scene in 2001), the identification process can be extraordinarily
difficult; a biological profile may be impossible; and in such cases, forensic scientists may rely heavily on comparisons with DNA reference samples, typically obtained
from relatives of the victims, to make positive identifications. To use DNA for identification, the scientist must have some knowledge of who the victim might have been
to find living relatives to whom DNA can be matched, or to find personal items such
as toothbrushes that might yield remnants of the victim’s own DNA. Without such
reference samples no identification can be made, although DNA may be able to narrow down the ancestry and identify the sex of the individual.
Besides DNA, other specific means for establishing a positive identification include
obtaining antemortem medical or dental records, examining surgical implants or
specific unique clothing or tattoos, or undertaking facial reconstruction (See Insights
and Advances: If You Have DNA, Why Bother with Bones? on pages 416–417).
Time Since Death
One of the more difficult tasks for a forensic anthropologist is determining how long
a victim has been dead. Anyone who has ever watched a police show knows that
body temperature can be used to estimate time since death if the death is sufficiently
recent. But over longer periods of time, other means are necessary. The research
program in forensic anthropology at the University of Tennessee maintains an outdoor morgue in which bodies are left to decompose under a variety of controlled
conditions so researchers can learn how natural processes affect the rate of decay
(Bass & Jefferson, 2003). Many other such programs are now being developed. All
the bodies used in the program are willed to the facility for this purpose, and once
they are skeletonized the remains are curated in a research collection for other types
of anthropological research including the development of comparative databases.
Decomposition is a continuum that includes a typical trajectory from cooling and
rigidity, to bloating, skin slippage, liquefaction, deflation, and skeletonization. The
rate at which decay proceeds is determined by aspects of the surrounding environment including burial depth, soil type, temperature, humidity, and so on. In general,
bodies left on the surface of the ground decompose most quickly and those buried
deeply in the ground most slowly. Surface remains decay more quickly because they
are more likely to be interfered with by scavengers, such as rodents and carnivores,
who destroy and scatter the remains. And insects also have greater access to surface
remains, speeding up decomposition. The timing of insect life cycles is well known,
and their preferences for certain types of tissues and extent of decay are also well
studied. Forensic entomologists therefore are important members of any forensic
team. Decomposition is quicker in the summer, averaging just a week or two for surface remains in the summer of the mid-Atlantic states. Although in very dry environments, such as deserts, bodies may mummify rather than skeletonize. The delay in
wintertime decomposition is due almost entirely to lower temperatures and humidity,
both of which reduce insect activity as well as the natural physiological rates of decay
of the body itself. Corpses that are wrapped in impervious containers, like garbage
bags, decompose more slowly than surface remains for many of the same reasons
that buried bodies decompose more slowly. With all things being equal, single burials tend to decompose more quickly than do the more protected individuals in the
middle of a mass grave—although individuals on the periphery decompose at rates
similar to those of individual burials. Using all these clues and others, scientists work
together to estimate time since death. This can help to narrow the focus of possible
identifications and possible perpetrators by suggesting a time frame for the crime.
Antemortem Records, Facial Reconstruction, and Positive IDs
Your dentist keeps a chart of which teeth you have, which have been extracted, and which
have been filled or crowned. All of your Xrays are also kept on file. These records can
prove invaluable for making positive identifications because no two mouths are the same.
However, comparing dental charts is time consuming, so the biological profile is used
to limit the scope of possible identities. Forensic odontologists, specially trained dentists,
work with forensic anthropologists to make identifications from dental records. Both dental Xrays and dental charts can be used for positive identifications (Adams, 2003). Exact
matches of antemortem and postmortem dental Xrays can establish an identification in
ways similar to antemortem medical Xrays (Stinson, 1975). But when Xrays are absent,
comparing dental charts is an effective means of identification as well. A dental chart is
made for the remains and this chart is compared, sometimes using the computerized
program, OdontoSearch, with antemortem charts of missing individuals (Adams, 2003) Medical Xrays taken before death can also be used for making identifications. An
Xray of a person’s head after an accident may reveal the frontal sinus, an air-filled space
within the frontal bone just behind the brow area (Figure 15.27). The sinus is uniquely
developed in each of us, so comparison of an Xray from a skull with an antemortem
film of a known individual may lead to a positive identification. Healed wounds and
infections that are caught on antemortem Xrays can also be compared to postmortem
Xrays. If the healing is particularly idiosyncratic, this might lead to a positive identification or at least to a possible identification that could be confirmed by other tests.
Orthopedic implants and pins often resolve issues of identity. These implanted
items often have either unique or batch serial numbers than can be traced back to an
individual patient’s medical records. And antemortem Xrays of a pin in place can also
be compared to postmortem Xrays to lead to a positive identification. Sometimes, the
biological profile doesn’t match any possible identities, and so there are no antemortem records to establish a positive ID. In these cases, other more exploratory methods,
such as facial reconstructions, may help the general public suggest a possible identity.
Facial reconstruction—the fleshing out of the skull to an approximation of
what the individual looked like in life—is part art and part science. It is based on
careful systematic studies of the relationship between skin thickness and bone features—and clay is used to layer on muscle, fat, and skin over a model of a victim’s
skull (Wilkinson, 2004). Digital technologies are also being developed to render
three-dimensional virtual reconstructions. Eyes and ears are placed, although their
color and shape can’t be known for sure. The size of the nose is based on the height
and breadth of the nasal aperture and the bony bridge. But some artistic license
is required to estimate the shape of the end of the nose. Skin and hair color can’t
be known from the bone. For example, facial reconstructions of King Tut were
commissioned by National Geographic magazine from two different artists. The two
yielded similar facial reconstructions, much of which were dictated by the king’s
uniquely shaped head and slightly asymmetric jaw, but inferences about weight,
skin and eye color varied. Once rendered, forensic facial reconstructions may be
photographed and shown to the general public in the hopes that someone might
recognize something about the individual. When possible identities are proposed,
antemortem records can be checked—and perhaps an ID will be made.
Applications of Forensic Anthropology
Although forensic anthropologists most often work on cases of lone victims of homicide, suicide, or accidental death, they are also called to the scene of mass fatalities,
to search for soldiers killed in combat, and to investigate human rights abuses that
result in hidden or mass graves.
Mass Fatalities
In the days after the attack on the World Trade Center in New York on September
11, 2001, forensic anthropologists from around the country were called in to help
identify the victims. The Oklahoma City bombing case seven years earlier had
brought in a similar influx of anthropologists, as did the later devastation wrought by
Hurricane Katrina. Forensic anthropologists play key roles in the attempt to identify
victims of earthquakes, plane crashes, floods, and other natural and human-wrought
disasters. The United States has regional emergency response teams called Disaster
Mortuary Teams (DMORT) that include pathologists, forensic anthropologists, and
forensic odontologists who are mobilized in response to national mass disasters such
as the World Trade Center fire and collapse. Military forensic experts and sometimes DMORT respond when U.S. citizens or military are involved in mass fatalities
abroad, such as the earthquake in Haiti for which DMORT was deployed to recover
remains of American citizens.
Although we often think of mass disasters as involving hundreds or thousands
of individual deaths, mass fatality incidents (MFIs) are defined as those in which the
number of deaths overwhelms local resources—there is thus no minimum number
and depending on the size of the municipality this might be fewer than 5 or 10
deaths. In such incidents, a main goal is to provide speedy and accurate disaster
victim identification (DVI), which requires three big operational areas—search and
recovery, morgue operations, and family assistance centers. Forensic anthropologists
are critical participants in all three of these areas.
In 2005, DMORT was deployed to assist in DVI in the aftermath of Hurricane
Katrina. Hurricane Katrina was a category 3 storm that devastated the Gulf Coast
of the United States in August of 2005. Up to that time it was the most costly storm
in U.S. history, and major damage was caused by the breaching of the levees that
protected the city of New Orleans, which sits below sea level. More than 1800 deaths
ensued, with flooding being the main cause of death and destruction. Because of
widespread flooding, victim recovery was also delayed, requiring the assistance of
forensic anthropologists in identifications. The identifications were made more
difficult by the flooding which also destroyed or damaged much of the antemortem
medical and dental records that are normally used in identification. DMORT teams
were rotated in for two-week assignments, and forensic anthropologist were utilized
in recovery, morgue operations, and family record collection.
War Dead
U.S. forensic anthropologists first became involved in the identification of those who
died in war when the Central Identification Laboratory (CIL) in Hawaii (CILHI) was
formed to aid in the identification of those missing in action during World War II.
Since then the skeletal remains of U.S. soldiers and civilians from World War II, the
Korean War, the Vietnam War, and other military actions have been recovered and
identified by this group of anthropologists. The CIL, now part of the Joint Prisoner
of War/Missing in Action Accounting Command (JPAC), sends teams around the
world to identify and recover U.S. soldiers lost in the wars of the twentieth century.
The remains are brought back to the CIL, thoroughly examined, and identified. In
addition to standard forensic anthropological techniques, JPAC teams also extensively use forensic DNA techniques to reach a positive identification so that remains
may be returned to the next of kin.
Not only does this group of forensic anthropologists help in identifying missing
personnel, but they have also undertaken some of the most important systematic research used in forensic anthropology and bioarchaeology. For example, Dr. Mildred
Trotter, an early director of the CIL, developed regression analyses for determining
stature from long bone lengths based on the skeletal remains of soldiers who died in
the Korean War. This large body of work remains a standard in forensic and bioarchaeological analyses today and would not have been possible without the detailed
medical histories of these military personnel.
War Crimes and Genocide
Finally, forensic anthropologists may play a key role in uncovering mass graves and
identifying bodies in them, and these scientists may be important witnesses in the
investigation of war crimes. Whether in Cambodia, Rwanda, Argentina, Bosnia, or
Iraq, when repressive regimes crack down on their citizens, they often attempt to intimidate the population through mass murder. The mass graves that are left contain
the bodies of hundreds or even thousands of victims, whose loved ones spend lifetimes attempting to locate them and determine their fate. Forensic anthropologists
help to identify the victims for the sake of surviving family members and may provide
key evidence in reconstructing a mass crime scene in an effort to bring those responsible to justice. Forensic anthropologists in these areas work for both government
and private groups such as Physicians for Human Rights, the International Commission for Missing Persons, and the United Nations (UN). Such teams often start work
before the conflicts end; for example, U.S. forensic archaeologists and anthropologists are currently at work in Iraq
One example of such work is the effort to exhume
mass graves in the former Yugoslavia that began in 1996
under the auspices of the United Nations, and in particular the International Criminal Tribunal for the former
Yugoslavia (ICTY), in partnership with Physicians for
Human Rights. Some of these exhumations concentrated
in eastern Croatia on a grave site known as Ovcara, which
contained victims from a massacre in Vukovar. The Vukovar
massacre occurred in November 1991. The forensic
teams exhumed about 200 bodies from Ovcara, nearly
all of them males (Figure 15.28). Mapping the grave site
took more than a month. The remains were autopsied in
Zagreb with the goals of constructing a biological profile
that would help in identification and interpreting perimortem trauma to understand the cause of death. Many
of the victims had multiple gunshot wounds and other
forms of perimortem trauma. Biological profiles were compared with the medical and dental records of missing people (a task hampered by
the destruction of hospitals and other medical facilities during the war) and lists
of identifying characteristics (including tattoos) provided by family members of
missing people. Through these comparisons, about half of the 200 were positively
identified. This evidence has been used in the prosecution of war crimes by the
UN-ICTY, including the case against the region’s former leader, Slobodan MiloŠevíc.
Forensic anthropologists use the changes wrought in the human skeleton by
natural selection and an individual’s life experiences to read the clues of recent human history. They apply the same principles, theory, and method to recent humans
that primate paleoanthropologists applied to understanding our 65-million-yearold fossil ancestors, all in a struggle to understand what makes us universally and
uniquely human.