By the end of the last Ice Age, humans
were living in most of the world’s
inhabitable places. They achieved a
global distribution without becoming
multiple species in the process, which
isn’t the way things usually h appen
in nature. They were able to do this
because they possessed extraordinary adaptive flexibility as biocultural
organisms. Without such flexibility,
humans might still be restricted to the
tropical and subtropical regions of the
Old World.
The archaeological record provides abundant evidence that the rate
of change in human lifeways accelerated markedly during the past 10,000
years or so. Throughout this long period, human culture accounted for more
and more of the changes in how and
where our ancestors lived. By making
cultural choices and devising new cultural solutions to age-old problems,
human groups succeeded in mitigating some of the processes that operate in the natural world (for example,
starvation due to seasonal food shortages) and either turned them to their
advantage or at least lessened their
worst effects. Humans also discovered that the cost of such cultural solutions included consequences for both
the natural world and human biology.
Chapter 16 will explore these consequences; some were fantastically good;
others weren’t.
Two of the most profound and farreaching developments of later prehistory were the shift from hunting and
gathering to food production and the
emergence of the early civilizations.
This chapter deals with plant and animal domestication and the associated
spread of farming, both of them integral to the development of the first
civilizations, which we will examine
in Chapter 15. We’ll start by examining competing explanations for the origins of food production. This will give
you a sense of the diverse perspectives from which researchers investigate how and why farming developed
after the end of the last Ice Age. From
there, we’ll discuss the archaeological
evidence for the origins of food production in several regions around the
world (Fig. 14-1).
The Neolithic Revolution
The change from hunting and gathering to agriculture is often called the
Neolithic revolution, a name coined
decades ago by archaeologist V. Gordon
Childe (1951) to acknowledge the fundamental changes brought about by the
beginnings of food production. While
hunter-gatherers collected whatever
foods nature made available, farmers
employed nature to produce only those
crops and animals that humans selected
for their own exclusive purposes.
Beyond domestication and farming, Neolithic activities had other farreaching consequences, including new
settlement patterns, new technologies,
and significant biocultural effects. The
emergence of food production eventually transformed most human societies either directly or indirectly and, in
the process, brought about dramatic
changes in the natural realm as well.
The world has been a very different
place ever since humans began developing agriculture.
Childe argued that maintaining fields
and herds demanded a long-term commitment from early farmers. Obliged to
stay in one area to oversee their crops,
Neolithic people became more or less
settled, or sedentary. As storable harvests gradually supported larger and
more permanent communities, towns
and cities developed in a few areas.
Within these larger settlements, fewer
people were directly involved in food
production, and new craft specializations emerged, such as cloth weaving,
pottery production, and metallurgy.
While archaeologists still accept his
general characterization of the Neolithic
revolution, we’ve learned a lot in the
decades since Childe’s original study.
For example, we now recognize that
sedentism actually preceded farming in certain locations where permanent settlements were sustained solely
by gathering and hunting or fishing. In
Chapter 13, for example, we noted that
the Chumash of southern California
and the Natufian hunter-gatherers of
the Near East, among others, established
sizable villages. It’s now also widely accepted that sedentism could often
stimulate food production, rather than
the other way around, and that even
Upper Paleolithic hunter-gatherers were
perfectly capable of understanding how
to manipulate the life histories of plants
and animals to their advantage.
Archaeologists also now know that
Neolithic lifeways evolved independently in several places around the
world. They are less agreed on how
and why it spread from one region to
another (e.g., compare Barker, 2006, and
Bellwood, 2005). Some researchers see
most regional Neolithic developments
as the culmination of local cultural
sequences. Others argue for migrations
and the diffusion of agriculture from “heartlands” in which such cultural
changes first took place.
Regardless of how it spread, the
Neolithic is viewed as revolutionary
in its cumulative impact on human
lives, not in the amount of time it took
for these cultural changes to happen.
Measured in human terms, the transition from hunter-gatherer to farmer in
any region involved complex, often interrelated biocultural changes that likely
played out across as many as 150 generations (Fuller, 2010, p. 11).
In the following discussion, we’ll
examine the beginnings of domestication and farming by considering evidence drawn from around the world.
Although the process everywhere shared
many similarities and produced equally
dramatic consequences, archaeologists
working in the Americas rarely apply the
term Neolithic to studies of New World
farmers. Instead, they use regional terminology, such as Formative or Preclassic
in Mesoamerica and Mississippian in
eastern North America.
Explaining the Origins of Domestication and Agriculture
Archaeologists have always felt compelled to identify “firsts.” When did
the earliest humans arrive in Australia?
Where are the oldest sites in South
America? When did the bow and arrow
arrive in the Midwest? Concerns about
these firsts also dominate archaeological research on domestication and agriculture and will undoubtedly continue
to do so. But we’ll never know when
or where the first person intentionally
planted seeds in the hope of making a
crop, and we’ll never track down the
first person to hitch an animal to a plow
or milk a goat. What researchers really
hope to achieve by their emphasis on
firsts is to understand what made these
changes happen.
From research on the origins of agriculture, we know that ancient huntergatherers who lived before the earliest
identified archaeological evidence of
food production were both intelligent
and observant enough to figure out what
happens to seeds after you put them in
the ground. In fact, we have every reason
to believe that Mesolithic/Epipaleolithic
and Archaic hunter-gatherers had a
wealth of practical everyday knowledge
and understanding about the natural
world around them. So, when we search
for the earliest evidence of agriculture in
a region, the most important goal is not
being able to say, “Ah, here’s where we
draw the line on our chronology chart of
agricultural beginnings” (see, for example, Fig. 14-1). A much more fundamental
motivation is simply to understand why
these people became farmers. Why then?
Why there? What conditions brought
about these changes? Why these crops?
From the perspective of the modern
world, we may find it hard to accept that
earlier peoples didn’t generally aspire
to be farmers and that many no doubt
avoided the opportunity for as long as
possible. But the archaeological record
and history alike are filled with examples demonstrating that what we now
view as the self-evident benefits of food
production have seldom been seen in
the same light by hunter-gatherers. In
recent centuries, many of the remaining hunter-gatherers on every continent
resisted, sometimes successfully, the
efforts of societies based on food production to convert them to peaceful, taxpaying farmers, voters, and consumers
of mass-produced goods. Such attitudes
call into question the inevitability of
agriculture in biocultural evolution. Did
it become the predominant economic
basis of human communities because it
offered so many obvious advantages?
Because it offered the fewest disadvantages? Or because there just weren’t a lot
of alternatives?
Ironically, although farming didn’t
get started in a big way until later, it was
the hunter-gatherers of the Mesolithic/
Epipaleolithic and Archaic periods who
actually initiated the critical processes
and even developed many of the innovations we usually credit to the Neolithic.
Essentially, the lifestyles of some early
Holocene hunter-gatherers anticipated
many of the developments we associate
with agriculture. Neolithic farmers were
mostly the recipients of domesticated species and agricultural ways from their
predecessors.
Defining Agriculture and Domestication
To avoid confusion later on, it’s useful to consider the difference between
domestication and agriculture. These
terms are often found together in discussions of the beginnings of food production, but they mean different things
(Rindos, 1984).
Domestication is an evolutionary
process. When we say that a certain plant
or animal is domesticated, we mean that
there’s interdependency between this
organism and humans, such that part of
its life history depends on human intervention. To achieve and maintain this
relationship requires the genetic transformation of a wild species by selective
breeding or other ways of interfering,
intentionally or not, with a species’ natural life processes.
Agriculture differs from domestication because it’s a cultural activity, not
an evolutionary process. It involves the
propagation and exploitation of domesticated plants and animals by humans.
Agriculture in its broad sense includes
all the activities associated with both
farming and animal herding. Although
domestication and agriculture are typically examined together in archaeological discussions of Neolithic lifeways,
domestication isn’t inevitably associated with an economic emphasis on food
production. For example, cotton was
an early domesticated plant, but it was
grown for its fibers, not as human food.
True agriculture would be unthinkable without domesticated plants and
animals. Domestication makes agriculture possible, especially when we consider how humans have manipulated
the life history strategies of other organisms to maximize particular qualities, such as yield per unit area, growth
rate to maturity, ease of processing,
seed color, average seed size, and flavor. The cultural activity we call agriculture ensures that the plants and animals with these desirable qualities are
predictably available as human food
and raw materials. One useful way
to view this fundamental change in
the relationship between humans and
other animals and plants is as symbiosis, a mutually beneficial association
between members of different species
(Rindos, 1984).
Next we’ll briefly examine some
competing explanations for the beginnings of agriculture. Although most of
these approaches address the problem of
explaining the development of farming
in the Near East, it’s important to note
that their proponents tend to view them
as generally applicable to agricultural
origins everywhere. The Near East dominates the discussion mostly because
it received the lion’s share of research
on this problem over the past century,
not because it was some sort of primal
hearth for the development of agriculture. Also, as you review these competing explanations, bear in mind that
the central questions are open areas of
research. Right now, no single approach
is both sufficient and necessary to
explain all known cases.
Loosely following Verhoeven (2004),
our overview examines two approaches,
those that primarily invoke natural, or
environmental, factors to explain the
development of agriculture, and those
largely based on cultural (including cognitive) factors.
Environmental Approaches
Most approaches to explain the origins
of domesticated plants and animals and
the beginnings of agriculture identify
one or more natural mechanisms, such
as climate change or human population growth, that may have promoted
the biocultural changes documented in
the archaeological record. The reasoning
behind such hypotheses is that, when
faced with increasing resource needs,
a community typically has several
options. The least disruption to everyday life can be achieved by reducing the
population, extending the territory, or
making more intensive use of the environment. Farming, of course, represents
a more intensive use of the environment.
Through their efforts, farmers attempt
to increase the land’s carrying capacity
by harnessing more of its energy for the
production of crops or animals that will
feed people (Fig. 14-2).
In their most extreme form, environmental approaches call to mind
environmental determinism, the notion that certain cultural outcomes can be
predicted from—or are determined
by—a combination of purely environmental causes. For example, V. Gordon
Childe himself conjectured that climate
changes at the end of the Pleistocene
increased Europe’s rainfall while making southwestern Asia and North Africa
much more arid (Childe, 1929, 1934).
Humans, animals, and vegetation in the
drought areas concentrated into shrinking zones around a few permanent
water sources. At these oases, Childe
hypothesized, the interaction between
humans and certain plants and animals
resulted in domestication of species such
as wheat, barley, sheep, and goats, which
people then began to use to their advantage. The eventual result was the spread
of sedentary village communities across
the Near East.
Hypotheses based on any form
of determinism tend to be relatively
straightforward, which is both their
strength and their weakness. Because
they hold so many factors constant, it’s
easy to see how such approaches should
work and why certain important outcomes should arise. The main drawback
of such ideas is that their focus is typically too general to explain a given case
because they omit the key contextual
factors that are unique to a real event.
In environmental approaches, such factors are often history and culture. What
people are already familiar with and
what they and their ancestors did in the
past often, if not always, play a big role
in their decisions. So, for example, a desert region might simultaneously sustain
opportunistic hunter-gatherers, nomadic
pastoralists, farmers using special deepplanting procedures, and even lawnmowing suburbanites willing to pay for
piped-in water, not because these groups
are unaware of the possibilities posed by
alternative ways of living, but because
they are living their traditional ways of
life and they prefer them.
To return to what has come to be
called Childe’s oasis theory, its simplicity quickly enabled archaeologist
Robert Braidwood to demonstrate that
the predicted outcomes didn’t exist in
the archaeological record (see p. 347 for
details). Pollen and sediment profiles
now confirm that at least some of the climatic changes hypothesized by Childe
did occur in parts of the Near East prior
to Neolithic times, and so they may have
had a role in fostering new relationships
between humans and other species in
this marginal environment (Henry, 1989;
Wright, 1993). Even so, both the causes
and the apparent effects were complex.
In places like the Near East, climate
change that resulted in diminished or
redistributed resources didn’t directly
push people to become farmers (Munro,
2004), though it may have made farming one of the more reasonable options.
Furthermore, the arid conditions familiar to us today in some parts of the Near
East may be as much a result as a cause
of Neolithic activities in the region (see,
e.g., Nentwig, 2007). That is, the ecologically disruptive activities of farmers
and herd animals during the Neolithic
period may have contributed to the
destructive process of desertification.
Their plowed fields exposed soil to wind
erosion and evaporation, while the irrigation demands of their crops lowered
the water table and increased salinization of the soil. And overgrazing herbivores rapidly reduced the vegetation
that holds moisture and binds soil, thus
destroying the fragile margin between
grassland and desert.
One large group of environmental
hypotheses that, unlike Childe’s, continues to be examined by researchers looks
to increased competition for resources.
Whether the competition resulted from
natural increases in population density
or from climatic changes, such as rising sea levels, increased rainfall, or lower 345
average seasonal temperatures, they’re
seen as major factors that encouraged
the domestication of plants and animals and, ultimately, the beginnings of
agriculture (e.g., Boserup, 1965; Binford,
1968; Flannery, 1973; Cohen, 1977). These
explanations share the view that agriculture developed in societies where competition for the resources necessary to
sustain life favored increasing the diversity of staple foods in the diet. For one
reason or another, population control or
territorial expansion may not have been
feasible or desirable choices in these
societies. For example, competition may
have arisen from decreased human mortality rates rather than increased fertility or possibly even been driven by the
increasing proportion of people living
to an old age. The point is that people
faced a “prehistoric food crisis” (Cohen,
1977) unlike most modern cases because
it was a chronic problem that worsened
over decades and showed no sign of
ever getting any better. Concentrated in
a restricted territory or faced with the
dwindling reliability of once-favored
resources, such hunter-gatherers might
have taken up horticulture or herding to
enhance the productivity or distribution
of one or more particularly useful species. It was this economic commitment
that eventually led to the emergence of
true farmers.
Binford’s (1968) “packing model”
develops one such hypothesis involving demographic stresses. As modern climatic conditions became established in the early Holocene, people
resided in every prime habitat in the
temperate regions of Eurasia. Foraging
areas became confined as territories
filled, leading to increased competition for resources and a more varied diet. Forced to make more intensive use of smaller segments of
habitat, hunter-gatherers applied their
Mesolithic technology to a broader
range of plant and animal species. A
few of these resources proved more
reliable, easier to catch or process, tastier, or even faster to reproduce than
others, so they soon received greater
attention. Archaeological evidence of
such changes can be found on many
Mesolithic/Epipaleolithic sites in the
form of sickles, baskets and other
containers, grinding slabs, and other
processing tools.
As local populations continued to
grow and other groups tried to expand
their territory, their only choice would
be to move into the marginal habitats that lay at the edges of the optimal, resource-rich parts of their territory (Binford, 1968). Because population
stress would quickly reach critical levels in these marginal environments,
where resources were already sparse,
it was here that domesticated plants
were first developed. To feed itself, the
expanding population might have tried
to expand the native ranges of some
of the resources they knew from their
homeland by sowing seeds of the wild
plants. Over time, this activity resulted
in domestication of those species and
fundamental changes in the relationship
between them and the people who by
then depended on them.
Aspects of Binford’s approach
appealed to many archaeologists, who
agreed that the wild ancestors of some
of the world’s most important domesticates originally held a low status in the
diet. Many were small, hard seeds that
were once seldom used except as secondary or emergency foods. Ethnographic
research had also shown that, given their
choice, hunter-gatherers everywhere prefer to eat fruit and meat (Yudkin, 1969).
Still, grains and roots became increasingly important in the Mesolithic/
Epipaleolithic diet—supplemented by
available animal or fish protein—and
not only when the more desirable foods
were in short supply. An interpretation based on increased competition for
resources offered a testable explanation
for why this happened.
Although he took issue with certain aspects of Binford’s approach,
Flannery (1973) agreed with the basic
thesis because it explained why the earliest archaeological evidence of plant
domestication should be found in what
would have been marginal environments. Flannery described the increasing breadth of the Epipaleolithic diet
as a “broad spectrum revolution” in
which hunter-gatherers turned to many
kinds of food resources to make up for
local shortfalls. Especially in marginal environments, this activity promoted the
development of domesticates and, ultimately, the origins of true agriculture.
In short, environmental approaches
identify forces external to humans as
the active ingredients in the development of agriculture. In these hypotheses, human agency is primarily reactive.
Something in the natural environment
changes (for example, precipitation patterns, average annual temperature),
and it makes life increasingly hard for
hunter-gatherers. They react culturally to
changed circumstances in various ways,
some of which include incorporating a
wider range of less preferred foods in
their staple diet and colonizing marginal
environments. At some point, they take
up the alternative of applying cultural
means to increase the production of one
or more food species. So basically, these
approaches envision the development
of agriculture more as something that
humans backed into from a lack of better
alternatives rather than something they
enthusiastically embraced.
Cultural Approaches
Not everyone agrees that the roots of
domestication and agriculture are to be
explained by the operation of external
environmental factors, all of which, by
design, place human culture and agency
in a passive role. Some archaeologists
contend that social and ideological factors, such as competitive feasting to
enhance one’s status, tribute payments,
or offerings to the deities (Price and BarYosef, 2010), may have pushed societies
to come up with more food than could
be readily obtained on a regular basis
from natural sources. The reasoning
behind these hypotheses is that human
agency and culture alone may be sufficient and necessary to explain many of
the fundamental changes documented in
the archaeological record.
As you may suspect from your
reading of the previous section, these
approaches are also not immune to
extreme positions. Just as we can identify some environmental approaches as
teetering on the brink of determinism,
we can find some cultural and cognitive approaches that seek to emphasize
the role of human culture to the near
exclusion of noncultural factors. In these
approaches, such natural phenomena
as climatic changes are either irrelevant
to the explanation of cultural outcomes
or were consciously exploited by people to further cultural objectives, so they
weren’t merely phenomena to which
people reacted.
Robert Braidwood’s “nuclear zone”
or “hilly flanks” hypothesis (Braidwood and Howe, 1960) is a good midtwentieth-century example to start
with in looking at cultural approaches.
Braidwood built on V. Gordon Childe’s
earlier work (see p. 344) and pointed
out that subsequent research didn’t find
evidence of the environmental changes
on which Childe based his oasis theory.
What’s more, the wild ancestors of
common domesticated plants and animals in the Near East were in the foothills of the mountains, not around the
oases, which is where they should be if
Childe’s oasis theory is correct. Without
a clear environmental trigger for the
origins of domestication and agriculture, Braidwood and Howe (1960) reasoned that domestication and, ultimately, agriculture came about as early
Holocene hunter-gatherers gradually
became familiar with local plant and
animal resources and grew increasingly
inclined to the notion of domestication.
In other words, domestication and agriculture happened when “culture was
ready.” But Braidwood never adequately
addressed the compelling questions that
such an argument stimulates: Why was
culture “ready”? Why then and not, say,
30,000 ya? Or 100,000 ya? Or never?
In his examination of the beginnings of agriculture in Europe, Ian
Hodder (1990) took a more evenhanded
approach than Braidwood. Building
on the symbolic meaning assigned to
houses and household activities, Hodder
developed an argument in which the
process of domestication and the activities of agriculture were properly viewed
as the human “transformation of nature
into culture, with an expansion of cultural control and a domination of
nature” (Verhoeven, 2004, p. 210). He
identified both social and natural factors
as possible pressures in bringing about
the transition from foraging to agriculture at the end of the Pleistocene. The
strength of Hodder’s approach rests in
his assignment of considerable weight
both to human agency and culture and
to the widely accepted effects of environmental factors. Trevor Watkins
(2010) makes a broadly similar claim.
Watkins argues that the key factor in the
“readiness” of human culture was the emergence of “larger and more cohesive
social groups” during the Epipaleolithic,
which stimulated human cognitive
development and, ultimately, the development of agriculture.
So, to sum things up, cultural
approaches to explain the origins of
domestication and agriculture assume
an active role for human agency and
tend to discount, if not deny completely,
the importance of natural, or environmental, factors. In these approaches,
cultural changes, such as a transformation of the human relationship with the
divine or of the conceptualization of self,
can be enough in some cases to account
for the changes we see in the archaeological record. The main drawback with
these hypotheses is that sometimes it’s
not immediately clear why such transformations would occur.
From Collecting to Cultivating
If today we had to choose an explanation for the origins of domestication and
agriculture (and we should at least suggest a preference, since this is an introductory college textbook), we would
adopt one of the moderate environmental approaches as the most robust
because it explains the most real cases.
Most such approaches also consider cultural factors, but they assign the greatest weight to the forces of nature. For
us, that’s their greatest appeal. They
don’t require researchers to assume that
just because we’re biocultural animals,
humans are somehow exempt from natural factors that affect all living things.
Disasters such as the devastating tsunami of December 2004 that killed more
than 180,000 people in a dozen countries
are painful reminders that, for all our
human posturing to the contrary, nature
often has the final word.
Ultimately, we have no reason to
believe that the origins of domestication and agriculture can be explained
only by natural forces or only by cultural
factors. These are complex problems for
which there may be multiple valid explanations. It could easily be the case that
approaches such as those recently proposed by Barker (2006) and Verhoeven
(2004), which seek explanations in the
interaction of both natural and cultural
forces, will prove to be the most productive route to follow.
So, working within our admitted preference for environmental
approaches, let’s now consider why and
how hunter-gatherers became farmers
in a real example drawn from the Near
East. As Epipaleolithic gatherers in the
Levant region harvested natural stands
of wild cereal grasses such as wheat or
barley, their movements would cause
many of the ripened seed heads to shatter spontaneously, with considerable
loss of grain. Each time someone used
a gazelle-horn sickle to cut through a
stalk, some of the seeds would fall to the
ground. This normal process of seed dispersal is a function of the rachis, a short
connector linking each seed to the primary stalk (Fig. 14-3). While the embryonic seed develops, the rachis serves as
an umbilical that conveys the nutrients
to be stored and later used by the germinating seed. Once the seed reaches
its full development on the stalk, the
rachis normally becomes dry and brittle,
enabling the seed to break away easily.
Even without human interference,
wild cereal grasses tended to be particularly susceptible to natural genetic
modification (much more so than, say,
nut-bearing trees), since the plants grew
together in dense patches, were highly
polytypic, and were quick to reproduce.
In fact, a stand of wild grasses was like
an enormous genetic laboratory. The normal range of genetic variability among
the grasses included some plants with
slightly larger seeds and others with
tougher or more flexible rachis segments,
meaning that their seed heads would be
slightly less prone to shattering. As people worked through the stands, seeds
from these genetic variants would end
up in the gathering baskets slightly more
often. Later, as the gatherers carried their
baskets to camp, stored or processed the
grain, or moved from place to place, a
disproportionate number of the seeds
they dropped, defecated, or perhaps
even scattered purposely in likely growing areas would carry the flexible-rachis
allele. (The same thing happened with
the larger seeds preferred by the collectors.) As these genetic variants became
isolated from the general wild population, each subsequent harvest advanced the “selection” process in favor of the
same desirable traits.
Human manipulation became an evolutionary force in modifying the species, a process Darwin labeled “unconscious selection.” People didn’t have to
be aware of genetic principles to act as
effective agents of evolution. And where
desirable traits could be readily discerned—larger grain size, plumper seed
heads, earlier maturity, and so forth—
human choice would even more predictably and consistently favor the preferred
characteristics. The result within just a
few growing seasons might be a significant shift in allele frequencies—that is,
evolution—resulting from Darwinian
selection processes, in this case the result
of long-term pressure by gatherers, who
consistently selected for those traits that
improved the plant’s productivity and
quality (Rindos, 1984).
Of course, the rate of divergent evolution away from the wild ancestral forms
of a plant (or animal) species accelerates
as people continue to exercise control
by selecting for genetically based characteristics they find desirable. With the
cereal grasses, such as barley and wheat,
the human-influenced varieties typically
came to average more grains per seed
head than their wild relatives had. The
rachis became less brittle in domesticated forms, making it easier for people to
harvest the grain with less loss because
the seed head no longer shattered to disperse its own seed. At the same time,
individual seed coats or husks (glumes)
became less tough, making them easier
for humans to process or digest. Many
of these changes obviously would have
been harmful to the plant under natural conditions. Frequently, a consequence
of domestication is that the plant species
becomes dependent on humans to disperse its seeds. After all, symbiosis is a
mutual, two-way relationship.
Whenever favorable plant traits developed, hunter-gatherers could be expected to respond to these improvements by
quickly adjusting their collecting behavior to take the greatest advantage, in
turn stimulating further genetic changes
in the subject plants and eventually producing a cultigen, or domesticate, under
human control. As continuous selection
and isolation from other plants of the
same species favored desirable genetic
variants, the steps to full domestication
would have been small ones.
Likewise, the distances that separated
hunter-gatherers from early farmers
were also small ones. It’s usually impossible to determine archaeologically when
harvesting activities may have expanded
to include the deliberate scattering of
selected wild seeds in new environments or the elimination of competing
plants by “weeding” or even burning
over a forest clearing. As they intensified
their focus on wheat and barley in the
Near East—or on species such as maize
or runner beans in Mexico—huntergatherers finally abandoned the rhythm
of their traditional food-collecting schedules and further committed themselves
to increasing the productivity of these
plants through cultivation.
Archaeological Evidence for Domestication and Agriculture
Archaeologist Graeme Barker (2006,
p. 414) argues that the change from
hunting and gathering to agriculture
“was the most profound revolution
in human history.” It also profoundly
affected other species and continues to
do so. The accumulated archaeological
evidence reveals that humans independently domesticated local species and
developed agriculture in several geographically separate regions relatively
soon after the Ice Age ended. From
recent applications of genetic research
and other methodological advances in
archaeology, it’s also clear that more
independent instances of prehistoric
domestication are likely to be identified,
some locations currently identified as
possible independent centers of domestication may be deleted from the list, and
some plants and animals were probably
domesticated multiple times in various
places (Armelagos and Harper, 2005).
Archaeologists have yet to fully
explain how and why agriculture
spread and eventually dominated economic life in many parts of the world.
Peter Bellwood’s (2005) recent “early farming dispersal hypothesis” integrates archaeology, genetics, and historical linguistics in a highly original argument that early farming grew out of the
Mesolithic/Epipaleolithic and spread
into new lands with the movement or
dispersal of farmers from agricultural
“homelands.” Working within what is
essentially an environmental approach
that identifies human population
growth as the main driving factor of
farmer dispersal, or “demic diffusion,”
this hypothesis offers a controversial
explanation for the spread of agriculture, human populations, and languages, as well as major spatial patterns
in human genetics (Bellwood, 2005;
Bellwood et al., 2007; see Diamond,
1999 for a very similar perspective).
Building on much the same archaeological and genetic data, Graeme Barker
(2006) reached different conclusions. He
essentially questions the existence
of such primal
hearths or homelands and traces
the roots of this
revolution back
to early Upper
Paleolithic huntergatherers. It is too
soon to tell which
of these interpretations offers a more
accurate picture of
the spread of prehistoric agriculture.
In examining
independent centers of domestication around the world, it’s important to
realize that the domestication of a local
species or two would not necessarily
trigger the enormous biocultural consequences we usually associate with the
Neolithic period in the Near East. In fact,
most altered species retained only local
significance. For example, in the Eastern
Woodlands of the United States, huntergatherers domesticated several smallseeded species very early; still, the wild
forest products obtained by hunting and
gathering retained their primary importance until relatively late prehistoric
times, when true farming developed in
the East.
In most regions, agriculture didn’t
develop fully until people were exploiting a mosaic of plants—and sometimes
animals, too—from different locations,
brought together in various combinations to meet such cultural requirements as nutrition, palatability, hardiness, yield, processing ease, and storage.
In the Near East, this threshold was
reached around 11,000 ya, when an agricultural complex consisting of wheat,
barley, sheep, and goats was widely and
rapidly adopted.
Plants
In most areas where agriculture
emerged, early farmers relied on local
plant species whose wild relatives grew
close by. Old World cereal grasses,
including barley and some wheat varieties, were native throughout the Near
East and perhaps into southeastern
Europe (Dennell, 1983). Wild varieties of these plants still flourish today
over parts of this range. Therefore, barley or wheat domestication could have
occurred anywhere in this region, possibly more than once. The same is true
for maize and beans in Mexico. So, as
we emphasized earlier, domestication
and agriculture were independently
invented in different regions around
the world.
As we noted earlier, it’s best to
explain these separate but parallel
processes from a combined cultural and
ecological perspective. We’ve already
seen that certain kinds of wild plants
were more likely than others to become
domesticated. Many of these species
tend to grow in regions where a very
long dry season follows a short wet period (Harlan, 1992). After the last Ice Age,
around 10,000 ya, these conditions existed around the Mediterranean basin and
the hilly areas of the Near East and in
the dry forests and savanna grasslands
of portions of sub-Saharan Africa, India,
southern California, southern Mexico,
and eastern and western South America.
Most scientific understanding of
ancient human plant use has come
from the archaeobotanical study of
preserved seeds, fruits, nutshell fragments, and other plant macrofossils like those shown in Figure 14-4 (Pearsall, 2000). It isn’t easy to preserve
seeds, tubers, leaves, and other delicate
organic materials for thousands of years.
Archaeologists recover some macrofossils from depositional environments that
are always dry, wet, or frozen, because
all of these conditions slow down or
halt the process of decomposition. Most,
however, are preserved because the
way they were harvested, threshed,
processed for consumption, or discarded
brought them into contact with enough
fire to char them, but not enough heat to
reduce them to ash. Once charred, macrofossils preserve well in many kinds
of archaeological sites and can often be
classified to genus, if not to species.
Macrofossils offer direct evidence
for important archaeological research,
such as reconstructing hunter- gatherer
plant use patterns, identifying farming locations, and determining the precise nature of harvested crops. They
also provide insights of other kinds.
The presence of perennial and biennial weed seeds in an ancient agricultural context may suggest that each year’s
farming activities only minimally disturbed the soil; the seed planter may
have used a digging stick, hoe, or simple scratch plow. If, on the other hand,
seeds of annual weeds predominate,
the farmer may have used a moldboard
plow—one that turns over the soil as it
cuts through it.
The major shortcomings of macrofossil-based interpretations of past
human plant use are due to potential
preservation biases. Most plant macrofossils tend to be preserved because they
were charred before entering the archaeological record (see Fig. 14-4 for examples). But not all plant parts char easily,
and many don’t char at all. For example,
it’s pretty easy to char a bean, but you’ll
be disappointed if you try the same
thing with a leaf of lettuce. Because the
necessary conditions could only be met
sometimes in prehistory and by some
plants and plant parts, archaeologists
are understandably concerned about the
validity and reliability of many reconstructions of human plant use that are
based only on macrofossils
Fortunately, modern archaeologists
can turn to several other important
sources in their research on prehistoric
human plant use. Plant
microfossils, such as
pollen, phytoliths, and
starch grains, often survive even where macrofossils can’t—for example, as residues on the
cutting edges of ancient
stone tools, inside pottery containers, embedded in the pores of
grinding stones, and
even trapped in the dental calculus that accumulates on human teeth
(Bryant, 2003; Henry
and Piperno, 2008).
Unlike many macrofossils, these remains preserve readily in a wide
range of archaeological contexts; they can be
classified as to the kind
of plant they represent,
if not also to the plant
part they represent; and
they can be archaeologically present even in
sites where macrofossils
were destroyed or never
deposited.
Pollen grains (Fig.
14-5) have been a valuable source of environmental and subsistence data for decades
(Traverse, 2007). Their
strengths are that they’re
abundant (as any hay fever sufferer can
tell you); the grains are taxonomically
distinctive and often can be classified to
genus, if not to species; the outer shell of
each grain is tough; and the wind-borne
dispersal of pollen from seed- producing
plants continues before, during, and
after humans occupy a particular
archaeological site. The main shortcoming of pollen grains is that they tend to
preserve poorly in many kinds of open
sites, depending on soil acidity, moisture, drainage, and weathering.
Phytoliths (Fig. 14-6) are microscopic,
inorganic structures that form in many
seed-producing plants as well as other
plants (Piperno, 2006). Like pollen, phytoliths are taxonomically distinctive.
They even vary according to where they form in the plant,
so phytoliths from
leaves can be distinguished from those
that formed in the
stems and seeds of
the same plant. They
are also affected by
different preservation biases than macrofossils (Cabanes
et al., 2011). Are the
plant remains at
your site unidentifiable, reduced to a
powdery ash, or simply not preserved?
Not a problem. Chances are the phytoliths from these plants are not only present in the archaeological deposit but also
recoverable and identifiable.
Starch grains (Fig. 14-7) are subcellular particles that form in all plant parts.
They are particularly abundant in such
economically important portions as
seeds and tubers (Coil et al., 2003). They
are a useful complement to phytoliths
as a data source and are a major tool
in archaeological investigations of root
crops (Piperno, 2008). Like pollen and
phytoliths, starch grains can be taxonomically classified, currently mostly to
family or genus. A good example of the
archaeological application of starch grain
analysis is the recent examination of the
surface of a grinding stone found on the
floor of one of the 23,000-year-old huts
at Ohalo II, in Israel. This study, which
was based on carefully sampled residues from cracks and pits in the working
surface of the grinding stone, enabled
archaeobotanists to identify that it was
a specialized implement used to grind
wild cereal grasses, including barley
(Piperno et al., 2004).
As we saw in Chapter 8, some plant
species also leave biochemical traces
in those who consume them. Because
temperate and tropical region plants
evolved with slightly different processes
for photosynthesis, their chemical compositions vary in the ratio of carbon-13
to carbon-12. This distinctive chemical
profile gets passed along the food chain,
and the bones of the human skeleton
may provide evidence of dietary change.
For example, their lower 13C levels reveal
that females at Grasshopper Pueblo, in
east-central Arizona, consumed mostly
the local plants they gathered, while
their male relatives at first enjoyed more
maize, a plant higher in 13C. Later, maize
became a staple in everyone’s diet at
Grasshopper, resulting in equivalent
carbon isotopes in males and females
(Ezzo, 1993).
Other biochemical analyses, using
different isotopes, have been devised
to assess overall diet—not necessarily
just the domesticated portions—from
individual skeletons. A higher ratio of
nitrogen-15 to nitrogen-14 (15N/14N), for
instance, corresponds to a greater seafood component to the diet (Schoeninger
et al., 1983), while a higher strontium-tocalcium (Sr/Ca) ratio indicates that plant
foods were of greater dietary importance than meat (Schoeninger, 1981).
Other chemicals taken up by bones may
inform us about ancient lifeways. For
example, lead (Pb) is a trace element
found in unusually high concentration in the bones of Romans who drank
wine stored in the lead containers typical of that period. The interplay among
culture, diet, and biology is, of course, a
prime example of biocultural evolution.
But to put it more simply, “You are what
you eat,” and the odds are increasingly
good that archaeologists can measure it.
Microfossil analyses complement and
greatly extend the valuable insights that
archaeobotanists have achieved through
the study of macrofossils. Recent key
advances include the growing field of
archaeogenetics, which applies the methods of molecular genetics to archaeological problems, as well as improvements in radiocarbon dating, which can
yield accurate age estimates from samples as small as 100 micrograms (μg)—
that’s one ten-thousandth of a gram, or
roughly one-fifth the weight of a grain of
rice (Armelagos and Harper, 2005). As a
result, researchers are now able to more
completely understand the prehistory of
the human use of plants and the beginnings of agriculture.
Animals
The process of animal domestication
differed from plant domestication, and
it probably varied even from one animal species to another. For example, the dog was one of the first domesticated
animals; mtDNA evidence suggests an
origin between 40,000 and 15,000 ya
(Savolainen, 2002), and dogs may even
have accompanied late Ice Age huntergatherers (Olsen, 1985). The dog’s relationship with humans was different
(and still is) from that of most subsequently domesticated animals. Often
valued less for its meat or hide, a
dog’s primary role was most likely as
a ferocious hunting weapon under at
least a bit of human control and direction. As people domesticated other animals, they changed the dog’s behavior even more for service as a herder
and later, in the Arctic and among the
Native Americans of the Great Plains,
as an occasional transporter of possessions. But the burial of a puppy with a
Natufian person who died some 12,000
ya in the Near East suggests that dogs
may have also earned a role as pets very
early (Davis and Valla, 1978).
Most other domesticated animals
were maintained solely for their meat at
first. Richard J. Harrison’s (1985) insightful analysis of faunal collections from
Neolithic sites in Spain and Portugal
concludes that meat remained the
primary product up until about 4,000 ya,
when subsequent changes in herd composition (age and sex ratios), slaughter patterns, and popularity of certain
breeds all point to new uses for some
livestock. Oxen pulled plows, horses carried people and things, cattle and goats
contributed milk products, and sheep
were raised for wool. Animal waste
became fertilizer in agricultural areas.
Leather, horn, and bone—and even
social status for the animals’ owners—
were other valued by-products.
Of course, animals are more mobile
than plants, and most of them are no less
mobile than the early people pursuing
them. So it’s unlikely that hunters could
have promoted useful genetic changes
in wild animals just by trying to restrict
their movements or by selective hunting alone. Possibly, by simultaneously
destroying wild predators and reducing the number of competing herbivores,
humans became surrogate protectors
of the herds, though this arrangement
would not have had the genetic impact
of actual domestication.
Since domestication is a process, not
an event, it’s nearly impossible to say
precisely when a plant or animal species has been domesticated. The process
involves much more than an indication
of “tameness” in the presence of humans.
More significant are the changes in
allele frequencies that result from selective breeding and isolation from wild
relatives. People may have started with
young animals spared by hunters or, in
the case of large and dangerous species
such as the auroch (the wild ancestor of
domesticated cattle), with individuals
that were exceptionally docile or small
(Fagan, 1993). Maintained in captivity,
these animals could be selectively bred
for desirable traits, such as more meat,
fat, wool, or strength. Once early farmers were consistently selecting breeding
stock according to certain criteria and
succeeding in perpetuating those characteristics through subsequent generations,
then domestication—that is, evolution—
clearly had begun.
Overall, not many mammal species
were ever domesticated. Those most
amenable to domestication are animals that form hierarchical herds, are
not likely to flee when frightened, and
are not strongly territorial (Diamond,
1989). In other words, animals that will
tolerate and transfer their allegiance
to human surrogates make the best
potential domesticates. Several large
Eurasian mammals met these specifications, and cultures of Asia, Europe, and
Africa came to rely on sheep and goats,
pigs, cattle, and horses (listed here in
approximately their order of domestication) as well as water buffalo, camels, reindeer, and a few other regionally
significant species.
Even fewer New World herd animals were capable of being domesticated. Aside from two South American
camelids—the llama and the alpaca—no
large American mammal was brought
fully under human control (Fig. 14-8).
Dogs had probably accompanied the
first people into the New World. None
of these animals were suitable for transporting or pulling heavy loads—llamas
balk at carrying more than about 100
pounds, and Plains Indian dogs dragged
only small bundles—so the people of the
New World continued to carry their own burdens, till their fields by hand, and
hunt and fight on foot until the introduction of the Old World’s livestock in
the 1500s.
Archaeological evidence of nonhuman animal domestication is subtle and
difficult to assess from the bones themselves (Fig. 14-9). Archaeozoologists,
who study the skeletal remains of nonhuman fauna from archaeological sites,
have also shown that
many of the morphological changes long
viewed as good proxy
measures of domestication (for example,
smaller adult size, horn
shape) actually follow,
by as much as thousands of years, other
archaeological measures
of domestication in the
Near East, such as distinctive demographic
patterns typical of managed herds (Zeder, 2008).
For example, at Ganj
Dareh, in Iran, around
9,900 ya, villagers
slaughtered a high proportion of young male
goats, but adult females survived long
after reaching reproductive maturity.
This culling pattern is typical of herded
animals (Zeder, 2008). If these villagers
were hunting wild goats, the demographic pattern evident in the goat bones
would have been different, emphasizing
large adults, mostly males.
Among important recent developments, DNA analyses of modern animals and ancient animal bones found in
archaeological sites are greatly enhancing our understanding of the domestication of individual animal species
as well as the development of farming
(Larson, 2011). Although not without
their own methodological and interpretive problems, population genetic analyses, coupled with new morphometric
analyses and recent advances in smallsample radiocarbon dating, hold considerable promise for answering some
of the big questions about the origins of
domestication.
Old World Farmers
As we’ve noted, independent invention
accounts best for the diversity of domesticates and the distinctiveness of Neolithic
lifeways around the world. What’s more,
many wild species, such as wheat and
sheep, probably occupied more extensive natural ranges in the early Holocene
and may in fact have undergone local
domestication more than a few times
(Armelagos and Harper, 2005).
But as Bellwood (2005) and Diamond
(1999) remind us, we can’t entirely discount the spread of Neolithic lifeways
or people from place to place. As we’ll
see, in at least some areas—southeastern
Europe, for example—colonizing farmers appear to have brought their domesticates and their culture with them as they
migrated into new territories in search
of suitable farmland. Neolithic practices
also spread through secondary contact
as people on the margins of established
farming societies acquired certain tools,
seeds, and ideas and passed them along
to cultures still further removed. Seeds
and animals must have become commodities in prehistoric exchange networks, just like the Spanish horses
obtained by Native Americans did in the
sixteenth century. It’s important to bear
in mind that each archaeological event is
unique in its own way. “One-size-fits-all”
explanations are rarely adequate, even if the results are the same—in this case,
the expansion of Neolithic lifeways. With
that in mind, let’s consider a sampling
of Neolithic societies from around the
world to get some idea of the variations
on this common theme.
The Near East
As we saw in Chapter 13, Epipaleolithic
foraging cultures, such as Kebaran and
Natufian, apparently took the first steps,
though perhaps inadvertently, toward
agriculture in the Near East (Fig. 14-10).
Many of the archaeologically documented changes among Epipaleolithic huntergatherers in the Levant offer important
support for Flannery’s (1973) conception
of a broad-spectrum revolution that preceded the development of domestication
and agriculture.
For example, the extraordinary preservation of plant remains at Ohalo II, an
early Epipaleolithic campsite in northern
Israel, shows that small-grained grass
and wild cereal seeds were important
in hunter-gatherer diets in the Levant
by 23,000 ya (see p. 352; Piperno et al.,
2004; Weiss et al., 2004a, 2004b). Faunal
remains from eight other Epipaleolithic
sites in the Levant, all of which date
between 18,000 and 12,000 ya, also show
a pattern of increased diet breadth as the
abundance of big game animals declined
and hunters depended more and more
on smaller game, such as gazelles (Stutz
et al., 2009).
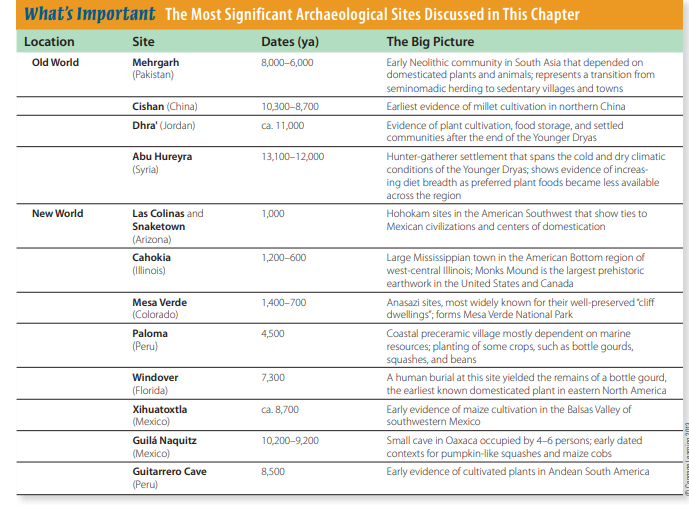
The site of Abu Hureyra, in the
upper Euphrates valley of Syria, is
of particular archaeological interest
because its occupation spans the
Younger Dryas (roughly 13,000–11,500
ya), a climatic shift to cooler and drier
conditions that David Henry (1989)
argues was an important environmental factor in the development of agriculture in the Levant. The Abu Hureyra
hunter-gatherers consumed more than
250 plant species, only a few of which
were staple foods, at the onset of the
Younger Dryas (Moore et al., 2000,
p. 397). As the climate changed, many
of these species vanished from the Abu
Hureyra archaeological record, to be
replaced by increased frequencies of
weed seeds and possible evidence
of early domesticated rye, specimens of
which date to roughly 11,200–10,600 ya.
Moore and colleagues (2010) infer
that changing environmental conditions were sufficient to account for
these shifts in Abu Hureyra’s subsistence economy and that this settlement depended at least partly on the
cultivation of domesticated plants (rye,
in this case). While the identification
of domesticated rye at Abu Hureyra
remains a controversial claim (see
Colledge and Conolly, 2010, pp. 135–
136), the subsistence economy clearly
reflects increased diet breadth as
higher-ranked foods became less available in the region around Abu Hureyra
(Colledge and Conolly, 2010).
Evidence of cultivation, settled communities, and food storage become
more common after the end of the
Younger Dryas and the return of a
slightly more moderate climate. At
Dhra’, an 11,000-year-old village site
near the Dead Sea in Jordan, archaeologists recently uncovered the remains
of mud granaries and other food storage features, as well as large quantities
of wild barley and oat grains (Kuijt and
Finlayson, 2009). The village economy
at Dhra’ clearly depended on the cultivation and storage of wild plants, and
many features traditionally associated
with sedentary Neolithic farmers are
seen at Dhra’.
Food collectors and the earliest farmers established the first permanent sedentary communities in the Near East.
Drawn by an ever-flowing spring in an
otherwise arid region, settlers at the
site of Jericho (or Tell es-Sultan), in the
West Bank, and at other Natufian sites
in Israel built their stone or mud-brick
round houses some 11,500 ya. Though
they were made of more substantial materials, in form these structures
closely resembled the temporary huts
of the region’s earlier hunters and gatherers. Numerous grinding stones and
clay-lined storage pits found in these
communities testify to the economic
importance of cereal grains (Moore,
1985; Bar-Yosef, 1987).
Finally, around 10,000 ya at Kebara
and El Wad, both in Israel, stone-bladed
sickles aided the harvest of wild wheat
and barley (Henry, 1989). This technology, more efficient than plucking seeds
by hand, netted greater yields during
a short harvest period. The process of
genetic selection that we’ve considered
as the basis for plant domestication must
have been well under way by this time.
Foraging for the seeds of wild cereals,
fruits, nuts, and the meat of wild game
long remained an important, but slowly
declining, strategy. By 10,000 ya, managed sheep and goat populations were
present in the Levant (Munro, 2009). At
Abu Hureyra, villagers were cultivating
wheat, rye, and lentils by 9,800 ya, but
wild plant food staples were still a part
of the diet (Moore et al., 2000). By 9,000
ya, sedentary villagers across a broad
arc from the Red Sea to western Iran—
also known as the Fertile Crescent (see
Fig. 14-10)—engaged in wheat and barley
agriculture and sheep and goat herding.
As demonstrated at many sites in this
region, Neolithic families lived in adjacent multiroom rectangular houses, in
contrast to the compounds of individual
small, round shelters commonly built by
Epipaleolithic collectors as well as the
early Jericho settlers.
Africa
Tracing Africa’s Neolithic past is challenging, considering the continent’s vast
size, its varied climates and vegetation
zones, and the extent to which many
of its regions are still archaeologically
unknown. What’s more, because many
tropical foods lack woody stems or durable seeds, plant macrofossils often are
poorly represented at African archaeological sites. Recent studies based on the
analysis of plant microfossils and genetic
patterns are helping to construct fresh
perspectives on the origins, development, and impacts of plant and animal
domestication throughout the African
continent.
Northern Africa Archaeologists working in the Nile Valley have found sickles
and milling stones relating to early
wild grain harvesting (Wendorf and
Schild, 1989). Qadan culture sites near
present-day Aswan (Fig. 14-11) probably
typified the Epipaleolithic food collectors who occupied the valley around
8,000 ya (Hoffman, 1991). Qadan people
employed spears or nets for taking large
Nile perch and catfish. Along the riverbanks they hunted wildfowl and gathered wild produce, processing starchy
aquatic tubers on their milling stones.
They also stalked the adjacent grasslands for gazelle and other game and
may have begun the process of domesticating the local wild cattle (Wendorf and
Schild, 1994; but see Gifford-Gonzalez
and Hanotte, 2011, p. 6). Considering the
wealth of naturally occurring resources
along the Nile, this foraging and collecting way of life might have continued
indefinitely. So why did farming develop
there at all?
Part of the answer rests with shifting rainfall patterns across North Africa
since the Late Pleistocene. Long-term
cycles brought increased precipitation,
which broadened the Nile and its valley and gave the river a predictable seasonal rhythm. Rains falling on its tropical headwaters, thousands of miles to the
south, caused the river to overflow its
downstream channels by late summer,
flooding the low-lying basins of northern Egypt for about three months of the
year. Although desiccation followed, the
flood-deposited silt grew lush with wild
grasses through the following season.
Periodically, however, extended drought
episodes intervened to narrow the river’s
life-giving flow.
West of the Nile, the fragile arid environment of the Sahara was particularly
susceptible to these fluctuations. It was
always marginal for humans, and down
to 11,000 ya, it was uninviting even
to hunter-gatherers. Then a period of
increased rainfall created shallow lakes
and streams that nurtured the Saharan
grasslands, attracting game animals and
humans. Around 7,000 ya, people in the
Sahara devised a strategy of nomadic
pastoralism, allowing their herds of
sheep, goats, and possibly cattle to act as
ecological intermediaries by converting
tough grasses to meat and by-products
useful to humans (Wendorf and Schild,
1994). Soon after, around 6,000 ya, further deterioration of the region’s climate—and possibly overgrazing—
forced the herders and their animals to
seek greener pastures closer to the Nile
and also to the south beyond the Sahara
(Williams, 1984; A. Smith, 1992; Kuper
and Kröpelin, 2006).
As drought parched the adjacent
areas, the Nile Valley attracted more settlers. Wild resources were then insufficient to feed the growing sedentary
population, and even the local domesticates that had been casually cultivated
on the floodplain gave way to more productive cereals—the domesticated barley
and wheat that had been brought under
human control elsewhere by people like
the Natufians.
Farmers gradually made the river’s
rhythm their own. Communities of
reed-mat or mud-brick houses appeared
across the Nile delta and along its banks.
Basket-lined storage pits or granaries,
milling stones, and sickles indicate a
heavy reliance on grain. Pottery vessels of river clay, linen woven from flax
fibers, flint tools, and occasional hammered copper items were produced
locally. These ordinary Neolithic beginnings laid the foundation for the remarkable Egyptian civilization, which we’ll
explore in Chapter 15.
Sub-Saharan Africa In West Africa
along the southern edge of the Sahara,
where little is currently known about the
early Holocene human presence, the pattern appears similar to that described for
the Sahara. At Ounjougou, in the Dogon
Plateau region of central Mali, erosional
gullies exposed a long sequence of
Pleistocene and early Holocene sites
(Huysecom et al., 2004). Excavations
reveal that hunter-gatherers lived in this region from the earliest Holocene,
around 12,000–11,000 ya. By 10,000–9,000
ya, descendants of these groups were
harvesting wild cereal grasses and making and using ceramics.
The major Near Eastern cereal crops
did not grow well in sub-Saharan Africa
or in its tropical regions. Between 5,000
and 3,000 ya, African farmers developed comparable domesticates from
local cereal grasses, including varieties of millet and sorghum (Fig. 14-12;
Smith, 1999). Pearl millet, an important food grain in parts of Africa and
India, was domesticated in Mali around
4,500 ya and was soon found across
sub-Saharan Africa. Manning and colleagues (2011) speculate that pearl millet may have spread rapidly because it
could grow even where there wasn’t
much water and it could be easily cultivated by mobile pastoralists. African
crops such as pearl millet, finger millet,
and sorghum found their way across the
Arabian Sea to South Asia more than
3,000 ya (Fuller et al., 2011), where they
are still raised today.
Hunters and gatherers in other parts
of Africa also experimented with local
cultivars. For example, mobile foragers and semisedentary fishers of tropical Africa practiced yam horticulture in
clearings and along riverbanks by 7,000–
6,000 ya (Ehret, 1984; Arnau et al, 2010).
With digging sticks, they pried out the
starchy tubers and carried them away
for cooking. As an added bonus, the
people discovered that if they pressed
the leafy tops or cuttings of the largest roots into the soil at the edge of the
camp clearing, the yams would regenerate into an informal garden. The domestication process for the yam, a tuber that
propagates vegetatively (for example, by
cuttings rather than by seeds), may have
taken far longer, possibly millennia, than
for the domestication of cereal grains
such as wheat.
More dramatic shifts in sub-Saharan
subsistence followed the introduction
and spread of several Neolithic domesticated plants and animals from South
and Southeast Asia across the Indian
Ocean. Among these economically
important plants, all of which spread
quickly throughout tropical Africa,
were bananas (or plantains) and two
root crops, taro and the greater yam
(Fuller et al., 2011, p. 550). The banana,
a food crop that is now of global importance, arrived in Africa from Southeast
Asia, possibly by 2,500 ya. Based on linguistic evidence, Blench (2009) argues
that plantains, taro, and the greater
yams were part of the Southeast Asian
“crop package,” which reached the
shores of Africa and quickly spread
throughout the interior.
Outside the Nile Valley, cattle herding took priority over farming in much
of East Africa, where conditions were
generally not suitable for cultivation.
Taurine cattle (Bos taurus) were first
domesticated in the Near East more
than 9,000 ya and by 8,000 ya could be
found on the savannas that then existed
in parts of the Sahara (Gifford-Gonzalez
and Hanotte, 2011). Genetic evidence also suggests a possible independent domestication of African taurine cattle (Bradley
and Magee, 2006). East African rock
paintings show that hump-backed zebu
(or indicine) cattle (Bos indicus), a South
Asian domesticate, reached Africa by
2,500 ya. The resulting African taurinezebu hybrid played a major role in the
success of East African cattle pastoralists
(Fuller et al., 2011).
Bantu-speaking peoples, native to
west-central Africa, relied on these and
other domesticated plants and animals
to support their rapid expansion through
central and southern Africa (Phillipson,
1984). Driving herds of domestic goats
and cattle and acquiring the technology
of ironworking as they moved southeastward through central Africa (Van
Noten and Raymaekers, 1987), the Bantu
easily overwhelmed most hunting and
gathering groups. The conventional
view is that with iron tools and weapons, they carved out gardens and maintained large semipermanent villages,
and today, their numerous descendants
live in eastern, southern, and southwestern Africa (see Fig. 14-11). There’s at
least some evidence that food- producing
methods spread into parts of southern
Africa before the Bantu. The bones of
domesticated sheep found in Late Stone
Age sites in South Africa may not be the
result of the spread of pastoralists, as
once widely believed, but the remains of
the camps of Bushmen (also called the
San), whom Karim Sadr (2003) describes
as “hunters-with-sheep.”
Asia
Several centers of domestication in
southern and eastern Asia gave rise
to separate Neolithic traditions based
on the propagation of productive local
plant and animal species. The exploitation of these resources spread widely
and heralded further economic and
social changes associated with the rise of
early civilizations in these regions (see
Chapter 15).
South Asia Excavations at Mehrgarh,
in central Pakistan, have illuminated
Neolithic beginnings on the Indian
subcontinent (Jarrige and Meadow,
1980; Fuller, 2006). Located at the edge
of a high plain west of the broad Indus
Valley (Fig. 14-13), the site’s
lower levels, dating between
8,000 and 6,000 ya, reveal the
trend toward dietary specialization that accompanied the
domestication of local plant and
animal species. Early on, the
people harvested both wild and
domesticated varieties of barley
and wheat, among other native
plants. Mehrgarh’s archaeological deposits also include bones
of many local herbivores: water
buffalo, gazelle, swamp deer,
goats, sheep, pigs, cattle, and
even elephants. By 6,000 ya, the
cultivated cereals prevailed,
along with just three animal
species—domestic sheep,
goats, and cattle. Researchers
believe that this early Neolithic
phase at Mehrgarh represents
a transition from seminomadic
herding to a more sedentary existence
that became the basis for later urban
development in the Indus Valley. Other
planned settlements boasting multiroom
mud-brick dwellings and granaries
soon appeared in the region, supported
by a productive agriculture and bustling trade in copper, turquoise, shells,
and cotton.
Rice is the staple crop of much of
South and East Asia. The earliest evidence of rice cultivation in India is
reported from the Lahuradewa site and
adjacent lake deposits in the Ganges
Valley, where cultivated rice phytoliths
and husks have been found in contexts
dated to around 8,360 ya (Saxena et al.,
2006). Domesticated rice spread from the
Ganges into South India after 3,000 ya,
possibly after the emergence of irrigation systems such as paddy-field cultivation (Fuller and Qin, 2009). The relationship between early South Asian
domesticated rice and that which was
domesticated by Chinese farmers in
the lower Yangtze Valley (see next section) is both unclear and an active area
of research.
Archaeologists know much less
about the origins of agriculture in South
India. Recent archaeobotanical analyses of samples taken from southern Neolithic, or Ash
Mound Tradition,
sites in Karnataka
and Andhra Pradesh
suggest that the earliest agriculture in
these regions dates
between 5,000 and
4,000 ya and was
based on several
native domesticated
species, principally
two lentils (horsegram and mung
bean) and two species of millet (browntop millet and bristly
foxtail grass), along
with nonnative crops
including wheat and
barley (Fuller et al.,
2004). The high frequencies of native
domesticates in these
samples lend support
to Vavilov’s (1992)
identification of India as a possible independent center of domestication.
China By 10,000 ya, hunter-gatherers
and East Asia’s earliest farmers were
already cultivating a crop in the Yellow
River basin that became a staple food
in the Neolithic villages and towns
of northern China. It may surprise
you to know that this crop was millet, not rice. The earliest firm evidence
of millet cultivation comes from the
early Neolithic site of Cishan, in Hebei
Province (Fig. 14-14), which has yielded
caches of common (or broomcorn) millet
in storage pits that date between 10,300
and 8,700 ya (Lu et al., 2009). Cishan was
part of a regional development of millet
farming that began with hunter- gatherer
cultivation in several places across
northern China. The deep stratigraphic
sequence at the Dadiwan site in Gansu
Province (Fig. 14-14) shows that the origins of millet farming can be traced to
the broad-spectrum resource strategy
of Late Pleistocene hunter-gatherers
in northern China (Barton et al., 2009;
Bettinger et al., 2010).
During the Yangshao period (about
7,000–5,000 ya), millet was a staple of
both humans and their domesticated
animals, which included pigs, chickens, and dogs, but for a long time foraging continued to provide wild plants,
fish, and animals. River terrace deposits of deep loess soil ensured large
yields and undoubtedly contributed to
the growth of populous settlements.
Domesticated rice is found in Yellow
River sites by the late Yangshao period, but in minor quantities relative to
millet; rice became fairly widespread
in this region after 5,000 ya (Fuller et
al., 2010)
At the Yangshao site of Banpo, in
Henan Province (see Fig. 14-14), more
than 100 pit houses were placed around
a plaza and its communal house, and a
protective ditch surrounded the village.
Cemeteries and pottery kilns typically
were located near the residential parts
of Yangshao villages. The jade carving,
painted ceramics of tripod form, silkworm cultivation, and elite burials found
in Yangshao sites anticipated some of
the hallmarks of the later Neolithic, or
Longshan, period beginning 4,700 ya.
Rice may have been cultivated by
hunter-gatherers in the lower Yangtze
region of southern China by 12,000–
10,000 ya (Fuller et al., 2010). Although
much is still unknown about the process
by which rice was eventually brought
into domestication, it was an economically important crop in the lower
Yangtze by 8,000 ya (Fuller et al., 2009;
Zhao, 2010). The population density of
the middle and lower Yangtze basin
increased rapidly between 7,000 and
5,500 ya, and substantial permanent villages were present throughout the region
(Zhang and Hung, 2008). Some of these
villages are considerably larger than
their neighbors and show evidence of
increased social complexity in their layout and structures.
Farmers introduced rice into
Southeast Asia over the next several
thousand years, bringing settled village
life and domesticated cattle, pigs, and
dogs to locations such as Ban Chiang
and Non Nok Tha, both in Thailand
(see Fig. 14-14). High yields and the varied conditions under which rice could
be grown made it the basis for sustained population growth in many parts
of this region (Higham and Lu, 1998;
Kharakwal et al., 2004)
Europe
Farmers in southeastern Europe already
were tilling the Balkan Peninsula by
about 9,000 ya at such sites as Argissa
and Franchthi Cave, in Greece (Fig. 14-15;
Perlès, 2001). In the eastern Mediterranean, Neolithic farmers arrived on
the island of Cyprus with domesticated
plants and animals from the Near East
as early as 10,000–9,000 ya (Zeder, 2008,
p. 11599). Far to the west, similar farmers reached the east coast of Spain
by 7,700 ya.
Researchers continue to debate
whether the spread of farming into
Europe was caused by the movement of
people, ideas, or some combination of
the two (Bellwood, 2005; Barker, 2006;
Colledge and Conolly, 2007; RowleyConwy, 2011). Considerable recent
archaeological, genetic, radiocarbon, and
other evidence supports the interpretation that the earliest farming cultures in
Europe were primarily the result of sporadic migrations by Neolithic farmers,
whose pathways stretched ultimately back to the Fertile Crescent. RowleyConwy (2011) and Zeder (2008) argue
that in many of the places successfully
colonized by these farmers, subsequent
interaction with indigenous huntergatherers facilitated the spread of farming, as well as domesticated plants and
animals, throughout parts of Europe.
This interpretation accounts for the
seemingly sudden appearance of fully
domesticated sheep, goats, wheat, and
barley at many sites in Europe and the
countries bordering the Mediterranean.
Had the Neolithic farming complex
diffused throughout Europe primarily through trade and exchange rather
than by migrations, farming would have
developed gradually in these regions
and the archaeological record would
yield evidence of “transitional” local
economies between hunting and gathering and farming. No such sites have yet
been found.
Neolithic lifeways transformed other
parts of Europe somewhat later than
southeastern Europe, and the source of
these changes has also generated debate.
Beginning around 7,000 ya, farming village sites littered with linear-decorated
pottery, or Bandkeramik (Fig. 14-16),
appeared across central and (still
later) northern regions of the continent (Bogucki, 1988). Bandkeramik culture farmers sought deep, well-drained
alluvial and loess soils located along the
Danube, the Rhine, and their tributaries. There they cultivated cereals and
legumes, raised cattle and pigs, and collected wild hazelnuts (Whittle, 1985;
Howell, 1987). Their settlements consisted of sturdy timber-framed structures
averaging 100 feet long, with some up
to 150 feet. These longhouses sheltered
extended families and possibly served
as barns for storing harvested crops or
for harboring animals. Wooden fences
barred livestock from planted fields during the growing season and then, following the harvest, confined them in the
field so that their manure could restore
soil nutrients. The fertility of loess soils
could be maintained for relatively long
periods with simple manuring and crop
rotation, and fixed-plot farming on rich
alluvium could sustain permanent settlements for up to 500 years (Whittle,
1985; Howell, 1987).
In Britain, Ireland, and southern
Scandinavia, the development of
Neolithic farming communities was
rapid and possibly traumatic (RowleyConwy, 2004). In the continuing debate
about whether European farming was
the product of indigenous development,
the cultural influence of farming neighbors, or migration from other regions, a
growing body of evidence makes a compelling argument for migration as the
main factor. For example, a recent analysis of radiocarbon dates from all regions
of Britain concludes that around 6,100–
5,400 ya, farmers from northern France
settled first in southwestern England
and shortly thereafter in central Scotland
(Collard et al., 2010). The results of this study are corroborated by the comparative analysis of pottery, faunal remains,
and mortuary practices and by stable isotope data that show a strong Mesolithic/
Neolithic diet shift (Richards et al., 2003;
Collard et al., 2010). Population density
also increased significantly across Britain
in the centuries following the arrival of
the island’s first farmers.
New World Farmers
While Old World Neolithic cultures
generally relied on agricultural practices that linked domesticated cereal
grasses together with herd animals,
the farmers of the Americas focused
almost exclusively on plant resources.
Most of these plants had very limited
ranges, but one important cereal grass—
maize, or corn—came to dominate prehistoric Native American agriculture
nearly everywhere it could be grown
(Fig. 14-17).
Cereal grasses suitable for domestication were abundant on several continents, but the people of the Old World
brought more of those species under control. Except for maize, whose wild ancestor may at first have been used in a different way, human cultural behavior in
the New World did not induce the same
genetic transformations in American
grasses as it had in the Old World.
New World Domesticates
Considered together, the products of
New World farmers make up a remarkable catalog of familiar plants, many
of which were domesticated between
10,000 and 7,000 ya (Iriarte, 2009).
Besides maize, the list includes important staple foods like white potatoes,
sweet potatoes, yams, manioc, many
bean varieties, peanuts, sunflowers,
and quinoa (Chenopodium sp.). Nearly
as important, but not staples, are such
domesticated vegetables and fruits as
sweet peppers, chili peppers, tomatoes, squashes, and pumpkins, along
with pineapples, papayas, avocados,
guavas, and passion fruit. Vanilla and
chocolate came from American tree
beans. Tobacco, coca, and peyote were
major stimulants, and a host of other
American plant domesticates had
medicinal, utilitarian, or ornamental uses long before the arrival of the
Europeans. The principal New World
domesticates were developed in several locations in Mexico and in South
America, but the use of a few of these
plants eventually spread far beyond
the regions in which they were first
domesticated.
One domesticated plant, the bottle
gourd (Lagenaria siceraria), appears to
have had a much different origin than
other New World domesticates. It was
long suspected that the bottle gourd
reached the New World by floating
across the Atlantic Ocean from Africa,
where it occurs in the wild, but genetic
research shows that the closest relatives of the oldest New World specimens
were from northeastern Asia. Based on
AMS radiocarbon dates of gourd samples and analyses of DNA markers, the domesticated bottle gourd was present
in the Americas by 10,000 ya and may
have been carried from Asia by PaleoIndian colonists (Erickson et al., 2005)!
Aside from the dog, which also
accompanied the first humans into the
New World, domesticated animals had
a relatively minor role in the Americas,
as we noted earlier (see pp. 353–354).
The llama and alpaca, both of which are
long-haired relatives of the camel and
are found in highland South America,
were the only large domesticated species
(Stahl, 2008). Other domesticates—the
guinea pig (raised for its meat, not as a
pet) and Muscovy duck in western South
America, turkeys in Mexico—were small
in size and not very important beyond
their localized distribution areas.
Mexico
Recent archaeological and genetic
research is forging a new understanding of the development of Mexican agriculture and of the beginnings of agriculture and settled farming communities in
the Americas. Of the more than 100 plant
species fully domesticated by Native
Americans in Mexico, maize (a grass),
beans (legumes), and squashes (cucurbits) ultimately attained the widest prehistoric significance for food purposes
(Iriarte, 2009).
Today’s many varieties of maize make
it one of the world’s primary staples (see
Fig. 14-17). Maize probably originated
somewhere in the valleys of southern
and western Mexico and was one of several plants domesticated during the early
Holocene. At the Xihuatoxtla site in the
Balsas Valley of southwestern Mexico
(Fig. 14-18), maize starch grains and
phytoliths have been recovered from
the surfaces of grinding stones and
other artifacts dating to about 8,700 ya
(Piperno et al., 2009; Ranere et al., 2009).
Similar evidence of early agriculture is
reported from the Gulf Coast lowlands
of Mexico, as well as at sites in Central
America and northern South America
(Iriarte, 2009).
But early Mexican agriculture was
not just all about maize. Other plants
were being brought under cultivation
in southern Mexico around the same
time, including several kinds of beans,
squashes, gourds, chili peppers, avocados, and cactus fruit (Flannery, 1986;
Smith, 1997). While long believed to
have been little more than a dietary
supplement for hunter-gatherers for
a long time, some of these crops were
actually capable of supporting communities. Others played important social
and ritual roles in Mesoamerica. The
best example of the latter is a tree crop,
cacao (the basic ingredient of chocolate).
It never became a major prehistoric
food crop, but it was ritually important
both as a beverage and in solid form.
Cacao use began in the Maya lowlands
between 3,900 and 3,700 ya (Powis et
al., 2007).
Maize, squash, and the common bean
make up an important trinity of prehistoric Native North American crops.
The common bean appears to have been
domesticated in west-central Mexico
(and again independently in Peru).
Comparative analysis of the genetic
diversity of wild and domesticated common beans points to the Lerma and
Santiago basin of western Mexico as the
region in which the common bean was
first domesticated in Mexico (Kwak et
al., 2009).
Like the common bean, squashes
were probably independently domesticated in several parts of the Americas.
In Mexico, domesticated seeds of
pumpkin-like squashes from Guilá
Naquitz Cave, in Oaxaca, are nearly
10,000 years old. Rinds of domesticated
squash have also been recovered from
8,700-year-old contexts at Xihuatoxtla, in
northern Guerrero (Ranere et al., 2009).
The maize-beans-squash crop trinity
eventually came together in the milpa
intercropping system that was an important part of Mexican agriculture.
South America
Given the analytical value of genetic
data and plant microfossils, research into
the history of domestication and agriculture in South America is making rapid
strides. The current focus is on several
major questions. First, what were the relative roles of marine resources and agricultural products throughout prehistory
on the continent’s west coast? Second,
to what extent did Mexican crops, particularly maize, contribute to South
American agriculture? Finally, what was
the nature of Amazonian agriculture in
eastern South America?
Sites in southwestern Ecuador have
yielded squash and gourd (cucurbit)
phytoliths that date to 12,000–10,000 ya.
Their large size suggests they’re from
domesticated plants; if this is true, it
means that the beginnings of food production in lowland South America began
about the same time as in Mesoamerica,
and maybe even earlier (Piperno and
Stothert, 2003).
Sediment cores and other geomorphological evidence indicate that the periodic climatic phenomenon known as El
Niño became established between 7,000
and 5,000 ya in the Pacific (Sandweiss et
al., 1996). El Niño events are triggered
when a persistent trough of atmospheric low pressure forces warm equatorial
waters southward along South America’s
west coast, partially displacing the
northward flow of deep cold currents.
El Niño typically disrupts the maritime
food chain and dramatically disturbs
precipitation patterns over land, bringing excess rainfall and flooding to some
areas, drought to others. El Niño returns
every four years or so, on average, and
some episodes are more severe or last
longer than others.
Early farming in coastal Peru seems
somewhat related to the El Niño pattern
(Piperno and Pearsall, 1998). At Paloma see Fig. 14-18), a short distance south
of present-day Lima, summer fishing
expeditions had extended into yearround reliance on large and small fish
species, shellfish, sea mammals, turtles,
and seabirds. Midden contents, analyses of coprolites, and high strontium
levels in human skeletons confirm the
nearly exclusive role of sea resources at
some sites by 5,000 ya (Moseley, 1992).
The fishers also began experimenting
with nonlocal plant crops, using bottle
gourds for carrying water and adding
several kinds of squashes and beans to
their diet. By about 4,500 ya, they had
taken up small-scale horticulture in
nearby river valleys, growing cotton for
nets and cloth and, significantly, supplementing their predominantly seafood
diet with at least 10 more edible plants.
While they maintained their basic maritime focus for centuries to come, coastal
Peruvians may have decided that a
greater variety of foods helped to minimize the periodic shortfalls in sea
resources that they could expect with
most El Niño events every few years.
The intercontinental dispersal or
exchange of American cultigens is a
topic of active archaeobotanical research
and debate (Smith, 1999; Iriarte, 2009).
Maize may have reached coastal South
America not long after being domesticated in southern Mexico (see Fig. 14-18).
Genetic data, preserved plant macroand microfossils, maize motifs on pottery, and even the impression of a kernel in the wall of a fired clay vessel, as
well as increases in both grinding stones
and human dental caries, attest to the
early presence of maize in this region.
Maize eventually became a significant
food source for all the native cultures
of western South America. Sixteenthcentury Spanish chroniclers noted that
many different varieties of maize accommodated Peru’s demanding climatic and
topographical diversity from sea level
to 6,500 feet, with potatoes taking over
at higher elevations. Each of these varieties, developed through careful selection and hybridization, probably derived
from a common ancestral form of
Mexican maize.
Plant cultivation was under way in
a few highland areas of South America
before 8,500 ya. Nonfood species useful for fiber, containers, tool shafts, bedding, and medicines were tended even
more often than edible plants around
Guitarrero Cave (see Fig. 14-18), in the
Andes Mountains (Lynch, 1980). Native
tree fruits, broad lima beans, smallseeded quinoa, and several starchy
tubers were among the local food crops
grown there (Lynch, 1980). One of these
ancient root crops was the white potato,
and when eventually adopted into Old
World agriculture and cuisine, it became
today’s familiar baked potato, Dutch
frites met, and Indian alu masala.
Other native South American cultigens were developed in the tropical forests on the eastern slopes of the Andes
or in the humid Amazon basin to the
east. Roots of manioc shrubs and sweet
potatoes became dietary staples in the
eastern lowlands, supplying abundant carbohydrate energy but little else.
Peanuts added some protein and fats
to the starchy diet, but fish and insects
remained essential food resources for
most of the natives of Amazonia, since
their small gardens alone generally
couldn’t sustain them entirely.
Southwestern United States
Agriculture based on maize and
other crops spread into the American
Southwest from Mesoamerica. The big
questions are currently why, when, and
how this northern dispersal of agriculture took place. Researchers are currently examining two possible explanations (Merrill et al., 2009). One view is
that maize agriculture was brought to
the Southwest by farmers who migrated
northward in search of suitable planting areas. Peter Bellwood (1997, 2005,
pp. 240–244) and Jane Hill (2001, 2010)
argue that these farmers were ProtoUto-Aztecan speakers, who began to
spread from their Mesoamerican homeland across the region around 3,500 ya.
This hypothesis, if supported by data,
would explain the diffusion of agriculture into the American Southwest, as
well as the subsequent distribution of
the Uto-Aztecan language family.
A competing hypothesis questions
the timing and role of the spread of the
Proto-Uto-Aztecan language family and
argues that maize agriculture entered the Southwest from Mesoamerica as the
result of “group-to-group diffusion,”
not migration (Merrill et al., 2009, 2010).
The proponents of this view argue that
Proto-Uto-Aztecan speakers were more
likely to have been Great Basin foragers, not Mesoamerican farmers, and that
the breakup of the Proto-Uto-Aztecan
speech community actually began in
the Great Basin nearly 9,000 ya. Merrill
and colleagues (2009) further note that
mtDNA research does not support a
Mesoamerican homeland for the ProtoUto-Aztecan language family (also see
Kemp et al., 2005).
The weight of evidence currently
supports the “group-to-group” diffusion hypothesis. The earliest maize is
present in the American Southwest
around 4,100 ya (Merrill et al., 2009).
Notably, other important Mesoamerican
cultigens, such as squash, amaranth,
beans, and cotton, did not arrive in
the region at the same time as part of
a coherent agricultural complex, but
reached the Southwest separately over
the course of the next two millennia.
Between 2,300 and 1,300 ya, reliance
on these domesticated products promoted increased population density
and overall cultural elaboration, resulting in the emergence of several distinctive prehistoric cultural traditions in the
Southwest (Fig. 14-19). Archaeologists
distinguish each of these regional traditions based on such features as pottery
style, architecture, religious ideas, and
sociopolitical organization (Plog, 1997).
The Hohokam of southern Arizona
were growing both food and cotton
by 1,500 ya and possibly much earlier,
irrigating their gardens through an
extensive system of hand-dug channels that conveyed water from the Gila
River or its tributaries. By around 1,000
ya, architecture and artifacts on large
Hohokam sites, such as Las Colinas
and Snaketown, near Phoenix, reveal
links to Mexican centers to the south.
They include ball courts and platform mounds as well as copper bells, parrot
feathers, and other Mesoamerican products (Haury, 1976). The Hohokam crafted
human figurines and shell and turquoise
ornaments in sufficient quantities for
trade (Crown, 1991). The Casas Grandes
district of northern Chihuahua, Mexico,
may have served as a major exchange
corridor between Mesoamerica and the
Southwest (DiPeso, 1974). But whether
the Hohokam maintained direct contact with Mexican civilizations or simply
participated in the diffusion of ideas and
products passed along trade routes is a
matter for debate.
The Mogollon, whose prehistoric culture straddled southern New Mexico
and Arizona, lived in pit houses until
about a.d. 1000. Around that time, they
began constructing aboveground room
blocks and—in imitation of their northern neighbors—creating boldly painted
black-on-white pottery. Archaeological
traces of the Mogollon faded a century
or more before Europeans arrived in the
mid-1500s, possibly as its people drifted
southward into Mexico.
In the Four Corners region to the
north, prehistoric farmers known to
archaeologists as the Anasazi built
impressive prehistoric masonry villages
and towns, called pueblos, beginning
around a.d. 900. With their large scale,
picturesque settings, and excellent preservation, some Anasazi sites— including
Chaco Canyon (Fig. 14-20), in New
Mexico, and the so-called cliff dwellings
of Mesa Verde (Fig. 14-21), in Colorado—
are among the most famous archaeological locations in the United States.
Anasazi towns consisted of multiroom,
multistory residential and storage structures and usually included underground
ceremonial chambers, called kivas. Their
compact sites were situated with good
access to the limited agricultural lands
and scarce water supply of this high and
arid region.
The rise of the Chaco Canyon towns
and related villages was probably stimulated by a brief period of increased
rainfall and sustained by social factors
such as political or religious ideology,
trade, and regional strife. A growing
body of evidence also points to Chacoera warfare and terrorism among the
Anasazi, extending even to cannibalism (White, 1992; Turner and Turner,
1999). Beginning in the mid-1100s, the
Anasazi abandoned Chaco and, eventually, most of the region’s other large
pueblos. By then, shifting precipitation patterns associated with a general
warming period were leaving marginal
zones of the Southwest, especially the
Colorado Plateau, without adequate rainfall to grow maize (Cordell, 1998). As
the drought worsened through the late
1200s, Anasazi townspeople persisted
in a few places like Mesa Verde (see Fig.
14-21), where they built their communities into easily defended niches in the
steep cliffs and tilled their fields on the
canyon rim by day. By a.d. 1300, even
Mesa Verde stood empty; the Anasazi of
the Four Corners had dispersed toward
the south and southeast to become the
people known today as Hopi, Zuni, and
the Rio Grande Puebloans.
Eastern North America
The earliest domesticated plant in eastern North America was the bottle
gourd, a specimen of which has been
found with a 7,300-year-old burial at
Windover, Florida (Doran et al., 1990).
Bottle gourds, which were grown for use
as containers, not as food, were probably domesticated in the Old World and
entered North America with the earliest
human inhabitants.
During the early Holocene, aboriginal
peoples developed an independent center of domestication and cultivation in
eastern North America (Price, 2009). The
first domesticated plant other than the
bottle gourd was squash about 5,025 ya,
followed soon by sunflower and marsh
elder (sumpweed). These plants are associated with ancient campsites and shell
heaps along major river valleys, where,
about 4,000 ya, people maintained “incidental gardens” of plants that supplemented traditional foraging activities
(Smith, 1999; Smith and Yarnell, 2009).
Over the next millennium, several more
native species, such as knotweed, maygrass, and little barley, were added to
the cultivated plants of the region. Stone
hoes also began to appear on sites in the
Illinois River valley at about the same
time (Odell, 1998).
It wasn’t easy to harvest and process
these weedy, small-seeded species, so
they probably weren’t much more than
supplements to a diet of wild foods. Still,
the river valleys of the Southeast and
the Midwest, as well as the rich forests
covering much of the Northeast, clearly
supported large, successful communities even without maize agriculture.
For example, the widespread practice of mound building and associated death
and burial rituals began long before any
significant reliance on maize. As we’ve
seen, Late Archaic hunter-gatherers also
constructed major mound groups that,
like those at Poverty Point, Louisiana
(see Chapter 13), were considerably more
than mere stacks of dirt.
Maize was not present in eastern
North America until around 2,150 ya,
and it appears to have had little economic impact at first. It’s not until after
1,200 ya in the Southeast and around 800
ya in the Northeast that we begin to see
archaeological evidence of a widespread
economic commitment to agriculture,
possibly brought about by new varieties
of maize and the introduction of domesticated beans (Smith, 1992; Hart and
Scarry, 1999). Even then, wild nuts, seeds,
fish, and game were staples in the diets
of many groups. In the broad river valleys of the Southeast, maize farming was
the economic mainstay of Mississippian
chiefdoms (Fig. 14-22).
Mississippian elites relied on elaborate rituals and displays of valued
symbols to enhance their privileged
positions. Populations and ceremonial
centers throughout the region were
linked by exchanges of symbolic copper,
shell, pottery, and stone items, as well as
a common focus on the construction of
towns centered on impressive earthenmound groups that flanked public
spaces or plazas (Lewis and Stout, 1998;
Emerson and Lewis, 2000). Most buildings that flanked the plazas were erected
on substructure or platform mounds.
Some of these buildings were dwellings; others were charnel houses and
other community structures. The houses
and workplaces of the town’s rank and
file clustered around these mound-andplaza complexes (Fig. 14-23).
The Mississippian site of Cahokia,
located below the junction of the
Missouri and Mississippi rivers near St.
Louis, once boasted some 120 mounds
(Milner, 1998; Pauketat and Wright,
2004). The primary earthwork was
Monks Mound (Fig. 14-24), as long as
three football fields and as high as a sixstory building—the largest prehistoric
structure north of Mexico. Cahokia’s
homes and garden plots spread over 6
square miles beyond the log stockade
that enclosed the central mounds and
elite living area. Fields of maize, squash,
and pumpkins extending along the river
floodplain provided the harvest sheltered in many storage pits and granaries
(Iseminger, 1996).
Other New World Regions
Elsewhere in the Americas, farming
hadn’t gained much importance even
by the time Europeans were arriving
with their own ways of life and their
Old World domesticates (Brown, 1994).
In fact, throughout much of western and
northern North America and in some
South American regions, hunting, fishing, and gathering were still the principal ways of making a living. These lifeways persisted in part because the
more productive American domesticates,
especially those native to warm temperate zones, couldn’t be introduced and
maintained in other geographical settings without sustained effort. Even so,
it wasn’t always a question of whether
farming was possible; it was often a
matter of choice. Maize was far from an
ideal crop, even where it could be grown
most readily. Old World domesticated
cereal grasses, including wheat, barley,
oat, rye, millet, and rice, grew in dense
stands that farmers could harvest readily
with a sickle and clean by threshing and
winnowing. Maize, the primary New
World cereal grass, required more space
per plant and more moisture during its
long growing season, and it was much
harder to harvest and process by hand.
What’s more, these agricultural products could seldom beat the nutritional
value of a mixed diet obtained through
foraging. Maize itself is deficient in
lysine (an amino acid) and niacin and
contains a chemical that may promote
iron-deficiency anemia. Similarly, in
the Amazon basin in South America,
the peoples who domesticated manioc,
sweet potatoes, and other starchy root
crops before 1,500 ya found that these
foods supplied bulk and carbohydrates
but little protein—a deficiency the people of the Amazon overcame by continuing to rely on hunting, fishing, and
gathering