I. Historical developments contributed for synthetic theory
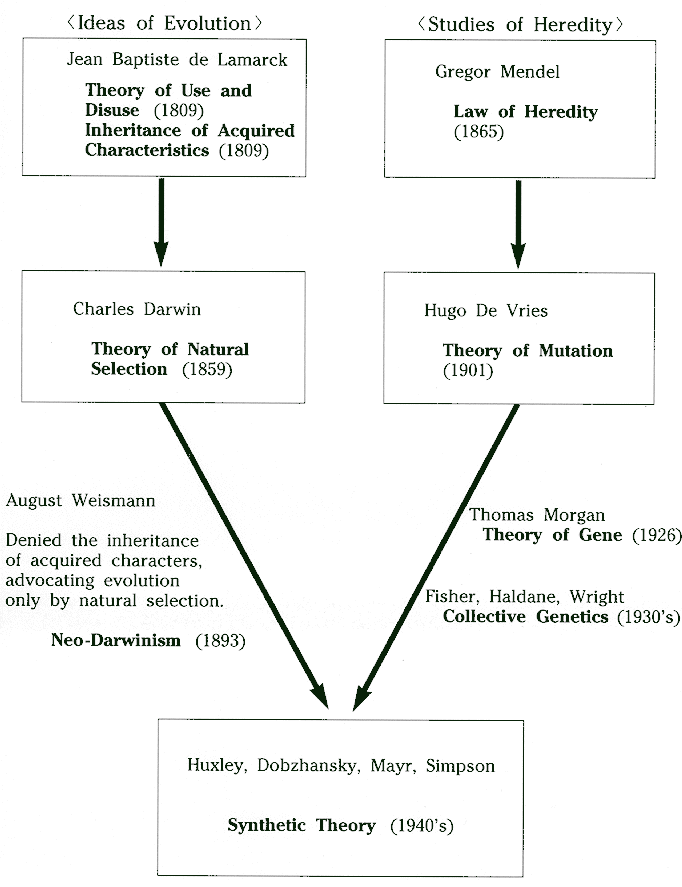
The idea of evolution is not entirely of recent origin. The essence of the idea appears in Greek writings of pre-Christian Era, such as those of Anaximander (600 BC), but it was never generally accepted. The scientific theory of evolution was not established until the eighteenth and nineteenth centuries. Major contributions towards developing and establishing evolutionary concepts were made by Lamarck and Darwin. During the post-Christian medieval period and until the 18th century, the Christian theory of special creation was the dominant thought. According to this theory, all living things come into existence in unchanging form due to Divine Will. The theory was notably in opposition to the concept of evolution. Another theory that received support during this period was the theory of spontaneous generation, which proposed that living organisms, at least the lower forms, arose spontaneously from non-living matter. The example of appearance of maggots in uncovered meat in a few days and tadpoles swimming in a puddle all of a sudden, were used to support spontaneous generation. However, experiments by Francesco Redi in 1668, Spallanzani in 1765 and finally Louis Pasteur in 1868 provided evidence that disproved this theory.
Contribution of Jean Baptist Lamarck: The first clear recognition and demonstration of the facts of evolution was made by the French naturalist Jean Baptist Lamarck (1774-1829). Lamarck discerned that all life is the product of evolutionary change, that evolution resulted in the taking on of new adaptations to the environment, and that the diversity of life was the result of adaptation.
Lamarck’s theory, popularly known as the “Theory of the Inheritance of Acquired Characters”, basically consisted of two parts.
- New structures appear because of an ‘inner want’ of the organism.
- These structures are acquired in response to need and are then inherited by subsequent generations.
He suggested that inner usage and need led to greater use or disuse of certain organs and tissues bringing about some modifications in the organism, which were passed on to the next generation. So an organism could pass on to its offspring any characteristic it had acquired in its lifetime. The elongation of neck from an original short-necked ancestor to the modern long-necked giraffe due to stretching to reach leaves higher up on trees, driven by ‘inner need’, is a common example of Lamarck’s theory. Lamarck was correct as far as the significance of evolution was concerned. But his theory of inheritance of acquired characters fails when subjected to scientific tests. Its failure was demonstrated by August Weismann (1834-1914), a German biologists, by cutting tails of mice for several generations. The offspring of tailless mice were always born with tails, thus experimentally disproving the inheritance of acquired characters. Though Lamarck was right in his insight as to the significance of evolution, but his theory does not stand up under investigation.
Contribution of Charles Darwin: The key theory of evolution was provided by genius of his time Charles R. Darwin (1809-1882), which converted the scientific and intellectual worlds to accept the fact of evolution. Darwin’s masterpiece book, “On the Origin of Species”, which appeared on November 24, 1859 had profound impact and changed our thinking forever in establishing the concept of evolution. Darwin’s theory is popularly known as the ‘Theory of Natural Selection’. Alfred Russell Wallace also came to similar conclusions independently and he communicated his thoughts about natural selection in a paper entitled “On the tendency of varieties to depart independently from the original type” to Darwin. Darwin communicated his own documents along with that of Wallace to the Linnaean Society and a joint presentation of two papers was made to the Linnaean Society on July 1, 1858. Their joint paper “On the tendency of Species to form Varieties; and on the Perpetuation of Varieties and Species by Natural Selection” was subsequently published in August 1858 in the ‘Journal of the Proceedings of the Linnaean Society’. This also prompted Darwin to publish in 1859 his theory of Natural Selection, which he had been developing for the last 20 years, in the form of a landmark book “On the Origin of Species”. According to Darwin, the prime force for evolutionary change was natural selection. Darwin’s theory of natural selection was based on two observations and two facts:
- All organisms reproduce many more offspring than can survive.
- Organism within a population exhibits variability and this variation affects an individual’s ability to survive and reproduce.
- The individuals best fitted to an environment survive, while individual variants less-suited fail to reproduce (natural selection).
- The traits (variations) thus favored by selection are passed on to the next generation .
Darwin gathered evidence for this theory during his voyage on the ship H.M.S Beagle. Darwin’s ideas were influenced by the work of economist Thomas Malthus (1766-1834). In his famous work, ‘An Assay on the Principal of Population’, Malthus explained how populations produce many more off springs than can possibly survive on the limited resources generally available. According to him, a fast growing population that reproduces too quickly can outstrip its resources and then crash (due to poverty, famine and disease) until the population size settles to a level which the resources can support. Darwin called this a “struggle for existence”.
Darwin realised that it was not chance alone that determined survival. Instead survival depended on the traits of an individual, which may aid or hinder survival and reproduction. Well-adapted or “fit” individuals are likely to leave more off springs than their less well-adapted competitors. Darwin realised that unequal ability of individuals to survive and reproduce could cause gradual change in a population. The characters (variations) that help an organism to survive and reproduce would accumulate in a population while those that hinder survival and reproduction would decrease or even disappear. Darwin used the term ‘Natural Selection’ to describe this process and perceived natural selection as the basic mechanism of evolution. Darwin correctly understood that natural selection is usually the most powerful mechanism of evolution but he did not completely comprehended how it operated .
One of the major weaknesses of Darwin’s theory was its inability to explain the sources of variations in traits within a species and how these variations were inherited. The hereditary mechanism (pangenesis) proposed by Darwin could not find wide acceptance. Darwin died before the mechanism of heredity was established in the late 19th and early 20th centuries.
Mutationism: ‘Mutationism’ refers to the theories of evolution where mutations are the main driving force of evolution. The concept of mutationism was first proposed in 1901 by Huge de Vries, the pioneer geneticist. Though later associated with Mendelian genetics, idea of mutationism began in the 1890s (before the rediscovery of Mendel’s laws) through the works of Hugo de Vries and Williams Bateson on naturally occurring discontinuous variations. This particular form of mutationism adopted by Hugo de Vries is often called ‘macromutation theory’ where suddenly large mutations could change radically a species into another.
The ‘mutationist view’ began by abandoning Darwin’s idea of automatic fluctuations, embracing instead the concept that variation emerges by rare events of mutations. This view was expressed in the writings of important founders of genetics, which included Thomas Hunt Morgan, Hugo de Vries and William Bateson, among others. Mutationists assumed that heritable variation could not be taken for granted. They understood evolution as a two-step process involving chance occurrence of a mutation, followed by its persistence or elimination. The mutationists denied that selection is creative and they conferred on mutation some extent of control over course of evolution. Unfortunately, early geneticists discarded all of Darwin’s ideas because they believed they knew something that Darwin did not know. It even became fashionable to talk of death of Darwinism during the first couple of decades of 20th century. Mutationist view was very popular in the first three decades of the 20th century but it was eventually replaced by the Darwinian view expressed in the ‘Modern Synthesis’ or ‘Neo-Darwinism’ or ‘Synthetic Theory’.
Hugo de Vries in the year 1900 proposed a new theory of evolution which is known as the Mutation Theory. This new theory did not consider natural selection as the principle force of evolution; rather it considered mutation as the main proponent of evolution. Mutations are called the spontaneous alteration of genes leading to changes in the organism and this in turn gives rise to new species. The new species originates suddenly and without any visible preparation.
The mutationists were of the opinion that most of the hereditary variables were discontinuous in nature and could be explained by the laws of Mendel. In this case, evolution would be effective if selection is operated on large mutations. Contrary to the mutationists, the biometricians led by Karl Pearson, supported the view of Darwin and argued that the major cause of evolution was natural selection. They opined that selection acting on small differences was the primary mechanism for evolutionary change. After the acceptance of the Hardy-Weinberg equilibrium (1908), mathematical models began to be developed and a new field called “population genetics” emerged. This field was developed largely due to the efforts of scientists such as Th. Dobzhansky, R. A. Fisher, S. Wright and J.B.S. Haldane.
Contribution of Mendel: For any discussion on the synthetic theory of evolution, the contribution of Mendel cannot be ignored. Darwin laid down the foundation of evolution by hypothesizing that if a trait is advantageous it will increase in frequency in a population because the offspring with the trait will survive and reproduce better and will pass on that trait to their offspring. However, at that time no one could explain how those traits could be passed over to the future generations. The credit for discovering the mode of inheritance of traits goes to the Austrian monk J. Gregor Mendel (1822-1884) who, through his breeding experiments on garden pea plants developed a few simple rules of inheritance. Mendel, the father of Modern Genetics, published his findings on inheritance in 1866 but his work remained largely ignored until it was rediscovered in 1900 by Hugo de Vries and Carl Correns. Mendel was a pioneer who laid the foundation for the whole of modern genetics. Rediscovery of Mendel’s principles led to the rapid and explosive growth of the discipline of genetics and established the basis for unraveling the deep secrets of biological reproduction and heredity. Mendel’s experiments helped him realise a few simple rules of inheritance.
Mendel proposed that there were discrete “factors” of heredity that united during fertilisation and then separated again in the formation of sperm and egg. It is remarkable how correct Mendel was, particularly in view of the fact that he knew nothing about DNA, chromosomes or meiosis (even the term ‘gene’ was not introduced until 1909). He was convinced that organisms inherit two units of each ‘factor’, one from each parent. Now we understand these ‘factors’ as genes and that most complex organisms are diploid, that is, they can have two copies of a particular gene or two different alleles and that alleles are not blended.
On the basis of his experiments, Mendel proposed the ‘Law of segregation’ . According to this law, when the gametes are formed in the parents, the heritable factors (genes) separate from each other so that each sperm or egg gets one unit of each pair. Mendel was correct. Today we understand that sperm and egg are haploid, with only half the number of chromosomes and genes of the parents. He also proposed what came to be known as the ‘Law of independent assortment’, which states that the factors (genes) for various traits assort independently of each other during the formation of sperm or egg. Mendel was partially right in this respect. It is because approximately 25,000 genes of the human genome do not float independently in the nucleus of the cell. Each gene is part of a homologous pair of chromosomes, and normally in humans there are only 23 pairs of chromosomes. Only genes meant for different traits, which are located on different chromosomes always truly assort independently.
Mendel published his findings in 1866, just seven years after Darwin’s ‘Origin of Species’. These findings went unnoticed until 1900, when eventually the mechanism of inheritance could be combined to natural selection. Shortly thereafter, a theoretical evolutionary model known as the ‘Modern Synthesis’ or ‘Synthetic Theory’ was born. Once in 1953, James Watson, Francis Crick and Rosalind Franklin explained the model of DNA molecule, the basic genetic component of evolution was revealed.
In other words, modern synthetic theory is basically about how evolution operates at the level of genes, phenotypes and populations.
II. SYNTHETIC THEORY
With the foregoing background of the basic concepts of evolution, let us now understand the synthetic theory of evolution. This theory of evolution is essentially a combination of Charles Darwin’s concept of natural selection, Gregor Mendel’s basic understanding of genetic inheritance, along with evolutionary theories developed since the early 20th century by field biologists, population geneticists, and more recently by molecular biologists. It is the present understanding of the process of evolution, which has been referred differently by different workers as ‘Neo-Darwinism, ‘Modern Synthesis’ and ‘Synthetic Theory’ etc. This is a combination of Darwinism selection with theoretical population genetics. The concept, also called as neo-Darwinism, was fundamentally developed by three founders of theoretical population genetics namely R.A. Fisher in 1930, Sewall Wright in 1931and J.B.S. Haldane in 1932 and later supported by others, especially Theodosius Dobzhansky. In fact, the scientists who were important in shaping the ‘modern synthesis’ theory included Theodosius Dobzhansky, Ernst Mary, R.A. Fisher and George Simpson. The book, ‘The Genetical Theory of Natural Selection’, by Fisher is considered a classic. The current understanding of the mechanism of evolution, though acknowledging the role of natural selection, differs considerably from the theory first outlined by Darwin. The advances in genetics, pioneered by Gregor Mendel, have led to a sophisticated understanding of the basis of variations and understanding of the basic of variations and the mechanism of inheritance. Researchers have identified DNA as the genetic material, through which traits are passed from parent to offspring, and identified genes as discrete elements within DNA. Though largely faithfully maintained within organism, DNA is both variable across individuals and subject to a process of change or mutation .
In Neo-Darwinism, natural selection is assumed to play a more important role than mutation, sometimes creating new characters in the presence of genetic recombination. Most geneticists believed that the amount of genetic variability contained in natural populations was so large that any genetic change could occur by natural selection without waiting for new mutations. Further mathematical geneticists showed that the gene frequency change by mutation was much smaller than the change by natural selection.
With a more complete understanding of mechanism of inheritance, the biological sciences now generally define evolution as the sum total of the genetically
inherited change in the individuals who are the members of the gene pool of a population. It is now understood that the effects of evolution are felt by the individuals but it is the population as a whole that actually evolves. Evolution, as per modern synthesis, can be expressed simply as a change in frequencies of
alleles in the gene pool of a population. The modern synthesis also emphasises the definition of species as a reproductively isolated group of organisms that
share a common gene pool . The modern theory of the mechanism of evolution differs from Darwinism in the following three significant
aspects:
- It recognises several forces of evolution in addition to natural selection, of which gene drift is equally important.
- It recognises that characteristics are inherited as discrete entities known as genes and that variation within a population is due to the presence of multiple alleles of a gene.
- It suggests that formation of new species is usually due to the gradual accumulation of small genetic changes.
In other words, modern synthetic theory is basically about how evolution operates at the level of genes, phenotypes and populations.
III. MECHANISM OF EVOLUTION
The synthetic theory explains evolution as consisting of two basic types of processes: those that introduce new genetic variations in a population, and those that affect the frequencies of variations already existing in a population. The raw materials for evolution are the variations. Without genetic variation the population cannot evolve.
Sources of variations
The various processes (or sources) that are responsible for introducing variations in a population are mutations and gene flow.
(a) Mutations
Mutations are considered as the ultimate source of all genetic variation. Broadly speaking mutation is alteration of genetic material. These are permanent, transmissible changes to the genetic material of a cell (usually DNA and RNA). These can be caused by ‘coping errors’ in the genetic material during cell division and also by exposure to radiation, chemicals or viruses. In multicellular organisms, the mutations can be divided in to ‘germline mutations’ (that occur in the gametes and can thus be passed on to the next generation) and ‘somatic mutations’ (which often lead to malfunction or death of a cell and can cause cancer). The mutations may be classified as ‘gene mutations’ and ‘chromosomal mutations’.
1) Gene Mutations: The spontaneous appearance of a new gene expression is called a gene mutation. It is the result of a slight change in the chemical structure of the segments of the DNA molecules that constitute a gene. It may be substitution, insertion or deletion of single base. For example, an adenine can be accidentally substituted for a guanine. Such errors in copying DNA are referred to as gene mutations or point mutations. Although there is self correcting mechanism in the replication of DNA that repairs such small errors but it may not always find and correct every one of them. Mutations can occur naturally as a result of occasional errors in DNA replication. Sometimes extra copies of one or more genes are produced when a DNA molecule is replicating. More often, however, sections of non-protein coding DNA regions are duplicated or inverted, which is an important source of genetic variation for a species. Spare copies of genes can mutate and change their function overtime thereby producing a new variation.
Mutations appear to be spontaneous in most instances. Point mutations may occur even in healthy people. Majority of them do not confer any significant advantage or disadvantage because they occur in non-gene coding regions of DNA molecules.
2) Chromosomal mutations: These are spontaneous changes in the structure or number of chromosomes. Chromosome mutations differ only in degree from gene mutations. The effect is more pronounced if the chromosomes undergo spontaneous modifications, than if only single gene mutates.
Chromosome mutations are inherited once they occur. These can be of various types given as below:
A) Change in number of chromosomes:
• 1) Gain or loss of a part of chromosomal set
• 2) Loss of an entire set of chromosomes (Haploidy)
• 3) Addition of one or more sets of chromosomes (Polyploidy)
B) Structural change in chromosomes:
1) Change in number of genes
• a) Loss of genes, usually termed as ‘deletion’
• b) Addition of genes, often called as ‘duplication’
2) Changes in arrangement of genes
• a) Exchange of parts between non-homologous pairs of chromosomes, termed as ‘translocation’
• b) Rotation of a group of genes by 180 degrees within one chromosome, often known as ‘inversion’.
Any of these major changes contributes to variability by changing the pattern of gene interaction. Structural modification of chromosomes generally occurs as a consequence of crossing over process during cell division. Normally, there is an equal exchange of end sections of homologous chromosomes. But sometimes, there is a reunion of an end section onto a chromosome that is not homologous. Similarly there can be an orphaned end section that does not reattach to any chromosome and the genes on such section are functionally lost. Irregular number of chromosomes can occur as a consequence of errors in meiosis and the combining of parental chromosomes at the time of conception . For example, there could be three instead of normal two autosomes for pair number 21, leading to a condition known as ‘Down Syndrome’ in humans.
For a mutation to be inherited, it must occur in the genetic material of a sex cell. Mutations provide genetic variability for various forces of evolution, such as natural selection, to operate upon. For a mutation to be subject to natural selection, it must be expressed in the phenotype of an individual. Selection favours mutations that result in adaptive phenotypes and eliminates non-adaptive ones. Even when mutations produce recessive alleles that are seldom expressed in phenotypes, they become part of a vast reservoir of hidden variability that can show up in future generations. Such potentially harmful recessive alleles add to the genetic load of a population, even mutations that have a natural effect could become advantageous or harmful if the environment changes to select for or against them.
The great diversity of life-forms seen in the fossil record is the evidence that there has been an accumulation of mutations producing a somewhat constant supply of variations upon which natural selection has operated for billions of years. Thus, mutation has been a significant prerequisite for the evolution of life. Another source of variation is recombination.
(b) Recombination
It is the process of mixing or recombining the existing genes into a variety of new genotypes. It is responsible for producing genetic combinations not found in earlier generations. Biologists recognise the importance of recombination as a source of genetic variation and as a significant partner with mutation, in manufacturing the materials for evolution.
Recombination does not produce new variations (or alleles) but only rearranges the existing genes in combinations not present earlier, thus adding to variability. New combinations of existing genes are produced at beginning of meiosis during crossing over when the ends of chromosomes break and reattach, usually on their homologous chromosome. This crossing-over process results in an unlinking and recombination of parental genes. Crossing over of the chromosomes is a phenomenon that only occurs during meiosis by chiasmata. Crossing over does not occur in asexual reproduction. During this process parental DNA gets exchanged and the resulting chromosomes contain a mosaic of genes from the mother and the father. The offspring has new combination of genes that did not exist in either of the parents. Crossing-over and recombination are crucial for maintaining genetic variability in successive generations. Recombination, thus adds greater diversity to the gene pool of a population. The recombination is significant to evolution. A single mutational change may be lost or passed without any significant effect on a population. However, its effect may be enhanced or modified by recombination. Thus variation is the raw material for evolutionary change, and gene recombination and mutation its principle source. Mutation alone may not have that pronounced effect without the impact of recombination in spreading it in a population.
Another source responsible for genetic variation is gene transfer. It is the movement of genetic material across species boundaries, including horizontal gene transfer, antigenic shift, and reassortment is considered as gene transfer. Viruses can transfer genes between species whereas bacteria can include genes from other dead bacteria or exchange genes with living bacteria thereby adding to dissimilarity.
Thus, variations are the raw materials on which various forces of evolution operate to effect evolutionary changes.
IV. GUIDING FORCES OF EVOLUTION
Natural selection and genetic drift are the major guiding forces of evolution that act upon the existing genetic variability within a species to cause evolutionary change.
(a) Natural Selection
It is one of the strong forces of evolution. Natural Selection as guiding force of evolution was the principle contribution of Charles Darwin to the evolutionary biology given as early as 1859 in his book “On the Origin of Species”. However Darwin did not fully comprehend how it operates, as mechanisms of genetics were not known at that time. Natural selection favored those features of an organism that brought it into a more efficient adaptive relationship with the environment. Through the process of natural selection, species become better adapted to their environments.
Under standard environmental conditions all genes in the gene pool of a population come to equilibrium, and this equilibrium is maintained. In other words, gene pool frequencies are inherently stable and do not change by themselves. The gene or allele frequencies will remain unaltered unless evolutionary mechanisms, such as natural selection and mutation, cause them to change. This understanding was largely developed through mathematical modeling based on probability in the early 20th century by an English mathematician Godfrey Hardy and a German physician Wilhelm Weinberg. Hardy and Weinberg postulated that gene frequencies in a population will remain stable if:
• all matings are random;
• mutation is not occurring;
• natural selection is not occurring;
• the population is infinitely large;
• all members of the population breed;
• there is no migration in or out of population;
• every one produces the same number of offspring.
That means evolution will not occur if no mechanisms of evolution are acting on a population, and the gene pool frequencies will remain unchanged. Hardy and Weinberg developed a simple equation that can be used to find out the probable genotype frequencies in a population and to track generational changes in them. This came to be known as the Hardy-Weinberg equilibrium equation (p2+ 2pq + q2= 1). In this equation, ‘p’ is defined as the frequency of dominant allele and ‘q’ as the frequency of the recessive allele for a trait controlled by a pair of alleles (A and a).
However, it is highly unlikely that any of these seven conditions, let alone all of them, is actually fulfilled in the actual world. Thus, evolution is inevitable. The natural processes that result in changes in gene pool from one generation to the next are the mechanisms that cause evolution. Natural selection is one such mechanism or force that brings about evolutionary change by favouring differential reproduction of genes, which produces change in gene frequency from one generation to the next. Natural selection on its own does not produce heritable genetic change, but once genetic change has occurred it acts to favour some genes over others. When two or more gene combinations are present, selection favours increased reproduction of the gene combinations most efficient under the environmental circumstances, thus bringing about improvement in adaptive relations between organisms and their environment.
Since the environment is never stable for greater period of time, the nature of selection process also fluctuates. In a changed environment a trait may lose its adaptive value and may not be encouraged by selection, which may start favouring another trait that improves ‘fitness’. It is probably that many of the slight changes in gene frequencies between generations may be due to changing selection pressures. The example of peppered moth (Biston betularia) in connection with industrial melanism in moths provides insight into the operation of selection under natural conditions. In England, until 1845, all known specimens of this moth were light in colour, but in that year a single black moth was taken at the growing industrial centre of Manchester. It is presumed that its highest frequency at that time in the moth population was not more than 1 percent. However, frequency of black moths increased to nearly 99 percent of Manchester moth population by 1895. This change in gene and genotype frequencies corresponded with the spread of industry in England. The change from light to dark color in dirty coal-dust covered areas of England is a good example of natural selection favouring dark pigmentation of moths, which provided better camouflage from predatory birds against dark background of soot-covered vegetation.
Natural selection moulds the genotypes of organisms so that they produce phenotypes best fitted to their environments. Consequently, the gene pool frequencies shift in the directions of their more adaptive alleles. But natural selection does not operate directly on the genotypes. It acts through the phenotypes of individuals and their gametes. It may be noted that whereas mutation and genetic drift are random, natural selection is not, as it preferentially selects for different mutations based on differential fitnesses.
(b) Sexual Selection
It occurs when organisms which are more attractive to the opposite sex, because of certain features, reproduce more and thus increase the frequency of those features in the gene pool. These features may not always have a conspicuous link to fitness. In humans, people usually select mates non-randomly for traits that are easily observable. Cultural values and social rules normally guide mate selection. Most commonly, mates are selected from among people who are like themselves phenotypically with respect to traits such as skin colour, stature and personality, etc. This is referred to as positive assortative mating. The net result of this type of mating is a progressive rise in the number of homozygous genotypes and equivalent decline in heterozygous ones in a population. Like recombination, non-random mating can act as an ancillary process for natural selection as a consequence evolutionary change. This is because any deviation from random mating upsets the equilibrium distribution of genotypes in a population thus affecting change in gene frequency or in other words causing evolution to occur.
However, the downside is that positive assortative mating results in an increase in homozygosity of deleterious alleles if they are present in the gene pool, particularly in some reproductively isolated small societies.
(c) Gene Flow
The phenomenon of transference of genes from one population to another is termed as gene flow. Evolution can occur due to gene flow as it can easily change gene pool frequencies even if no other mechanisms of evolution are operating. Gene flow takes place when there is migration of individuals into (immigrations) or out of (emigration) the population. This physical movement of alleles, called gene flow, tends to mix pools of genes that might not otherwise mingle. The prevalence of international travel now a days has markedly increased the possibility of gene flow in human population. Human gene flow is regulated to a certain extent by culture, which determines how frequently populations interact and interbreed. Gene flow mixes the alleles of different gene pools, thus stopping them from diverging into separate species. Genes may occasionally also flow between species. For example, segments of DNA may be transferred from one species to another by viruses as they invade cells of other organisms. This rare form of gene flow has been reported for some species of fish, reptiles, insects, mammals and microorganisms, but it has yet to be conclusively demonstrated for humans.
(d) Genetic Drift
Although natural selection is a significant natural force; but it is not the only force acting upon variation to produce evolutionary change. One such force is genetic drift. It refers to the random changes in allele frequency because of chance events. In small, reproductively isolated populations, chance factors produce rapid changes in gene frequencies totally independent of mutation, recombination, and natural selection. The smaller the populations, the more susceptible it is to such random changes. The genetic drift operates in both large and small populations, but it is only in the latter that these processes produce a significant evolutionary effect. Studies in population genetics, principally by Sewall Wright, have led to the recognition of the force of genetic drift or Sewall Wright effect that plays an important role in population evolution.
Genetic drift, usually referred to as drift in evolution, mainly occurs in small populations and may fix certain non-adaptive or neutral genes. The essential feature of genetic drift is that the smaller the population, the greater are the random variations in gene frequencies from one generation to the next. Thus, drift must have played a major role in the early stages of human populations which were numerically very small. However, even in populations of today, there
are culturally isolated groups, such as Jarwa and Onge of Andaman Islands of India in which drift can still be an important evolutionary mechanism.
One cause of genetic drift is the ‘founder effect’ or the ‘founder principle’. It occurs when a few individuals leave the original group and begin a new population in a different location, or when some environmental change isolates a small population from a larger one. The new gene pool may not be a representative of the original population. The resultant new population usually contains high frequencies of specific genetic traits inherited from the few common ancestors who first had them.
Another cause of genetic drift is the ‘bottleneck effect’, which occurs when a major catastrophe such as sudden climatic change, fire, devastating earthquake or a new predator wipes out most of a population without regard to any previous measure of fitness. When this happens, the few genes left in the gene pool may no longer represent the original population, nor do they necessarily represent the fit genes. The few survivors of such evolutionary bottlenecks may reproduce resulting in a large population in subsequent generations. The consequence of the bottleneck effect is the drastic reduction in genetic diversity of species since most variability is lost at the time of bottleneck.
Speciation
The evolutionary process of formation of two or more species from one is known as speciation. A species is basically a taxonomic classificatory category. There are several species concepts that use different criteria to define a species. The biological species concept is the most commonly used concept to group living animals. According to this concept, a species is a natural population in which individuals are actually or potentially capable of breeding with one another to produce fertile, viable offspring and, under natural conditions; do not normally interbreed with individuals of other species.
The accumulated microevolution or changes in allele frequencies in a population lead to macro-evolution, which is speciation. In a sense, variation that exists within populations increases to become variation between populations. Evolution at the species level is the outcome of cumulative microevolution. The accumulated changes may ultimately lead to reproductive incompatibility and thus to the creation of species. For speciation to occur, some form of isolating mechanism, even if only partial or spatial, is necessary for evolutionary divergence. Without isolation the gene flow precludes evolutionary divergence. With isolation, microevolution within the population groups (demes) may produce morbidly divergent populations. The formation of two or more species often requires geographical isolation of subpopulations of the species. Only then mechanisms of evolution such as natural selection or genetic drift produce distinctive gene pools. Isolation is therefore the key factor in the origin of new populations and ultimately new species. Populations of a species may get isolated due to geographic barriers like river and desert etc or by sheer distance in populations having large geographic distribution, or islands separated by rising sea levels to isolate subset of the population, or by mountain chains, etc. Depending upon the extent of geographic isolation, four different modes of speciation are commonly understood viz., allopatric speciation, parapetric speciation, paripetric speciation and sympatric speciation.
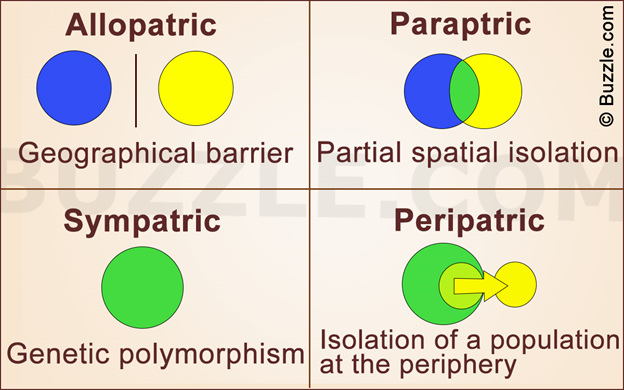
Allopatric speciation : occurs when a species separates into two separate groups which are isolated from one another. A physical barrier, such as a mountain range or a waterway, makes it impossible for them to breed with one another. Each species develops differently based on the demands of their unique habitat or the genetic characteristics of the group that are passed on to offspring. When Arizona’s Grand Canyon formed, squirrels and other small mammals that had once been part of a single population could no longer contact and reproduce with each other across this new geographic barrier. They could no longer interbreed. The squirrel population underwent allopatric speciation. Today, two separate squirrel species inhabit the north and south rims of the canyon. On the other hand, birds and other species that could easily cross this barrier continued to interbreed and were not divided into separate populations.
Peripatric speciation : When small groups of individuals break off from the larger group and form a new species, this is called peripatric speciation . As in allopatric speciation, physical barriers make it impossible for members of the groups to interbreed with one another. The main difference between allopatric speciation and peripatric speciation is that in peripatric speciation, one group is much smaller than the other. Unique characteristics of the smaller groups are passed on to future generations of the group, making those traits more common among that group and distinguishing it from the others.
Parapatric speciation : a species is spread out over a large geographic area. Although it is possible for any member of the species to mate with another member, individuals only mate with those in their own geographic region. Like allopatric and peripatric speciation, different habitats influence the development of different species in parapatric speciation. Instead of being separated by a physical barrier, the species are separated by differences in the same environment.
Parapatric speciation sometimes happens when part of an environment has been polluted. Mining activities leave waste with high amounts of metals like lead and zinc. These metals are absorbed into the soil, preventing most plants from growing. Some grasses, such as buffalo grass, can tolerate the metals. Buffalo grass, also known as vanilla grass, is native to Europe and Asia, but is now found throughout North and South America, too. Buffalo grass has become a unique species from the grasses that grow in areas not polluted by metals. Long distances can make it impractical to travel to reproduce with other members of the species. Buffalo grass seeds pass on the characteristics of the members in that region to offspring. Sometimes a species that is formed by parapatric speciation is especially suited to survive in a different kind of environment than the original species.
Sympatric speciation : is controversial. Some scientists don’t believe it exists. Sympatric speciation occurs when there are no physical barriers preventing any members of a species from mating with another, and all members are in close proximity to one another. A new species, perhaps based on a different food source or characteristic, seems to develop spontaneously. The theory is that some individuals become dependent on certain aspects of an environment—such as shelter or food sources—while others do not.
A possible example of sympatric speciation is the apple maggot, an insect that lays its eggs inside the fruit of an apple, causing it to rot. As the apple falls from the tree, the maggots dig in the ground before emerging as flies several months later. The apple maggot originally laid its eggs in the fruit of a relative of the apple—a fruit called a hawthorn. After apples were introduced to North America in the 19th century, a type of maggot developed that only lays its eggs in apples. The original hawthorn species still only lays its eggs in hawthorns. The two types of maggots are not different species yet, but many scientists believe they are undergoing the process of sympatric speciation.
V. NEO-MUTATIONISM
Mutationism, which began in the 1890 with the studies of Hugo de Vries and William Bateson, was very popular view in the first three decades of the 20th century. It was eventually replaced by the ‘Modern Synthesis’ or ‘Synthetic Theory’, which is currently a widely accepted theory of evolutionary mechanism. The contemporary view corresponding to mutationism could be termed as NeoMutationism. With the arrival of molecular biology, interest in mutations as an important factor in protein evolution has been shown by some scientists. A new view is emerging that uses recent molecular studies of phenotypic evolution to
support the basic ideas of mutationism put forth by Thomas Morgan in 1932. This is a new form of mutation theory on the role of mutation in the evolution and is often referred to as neo-mutationism. One of the strongest advocates of neo-mutationism is Masatoshi Nei. Scientists studying molecular evolution began to suggest mutational explanation for patterns such as genomic nucleotide composition. Phrases such as ‘new mutations’ or ‘mutation–driven evolution’
have emerged in the recent few decades that advocate a departure from the ‘changing gene frequencies’ view of ‘Modern Synthesis’ or ‘Synthetic Theory’. These contemporary workers suggest that mutation plays a role in evolution that was initially proposed by ‘mutationists’ but rejected by Modern Synthesis.
A main feature of this theory is how single mutations can have significant effects to influence evolution.
While neo-Darwinism regards mutation as merely raw material and natural selection as the creative force, the Neo-mutationism of Nei assumes that the most fundamental process for adaptive evolution is the production of fundamentally more efficient genotypes by mutation (especially birth and death process of duplicated genes) and the recombination. Neo-mutationism suggests that the basic genetic factor of phenotypic evolution is the mutational change of protein-coding regulatory regions of genes and that evolutionary change of phenotypic characters is caused by the so called ‘major-effect mutations’. It further assumes that natural selection occurs as a consequence of mutational production of different genotypes, and therefore it is not a cause of evolution. According to Nei’s view, mutation (including gene duplication and other DNA changes) is the driving force of evolution at levels of genes as well as phenotypes.