- Who were Homo erectus ?
- Variations and their Distribution
- Time Period
- Anatomical Features
- Cultural Remains
- Other Aspects of Culture
- Dispersal of Homo erectus
- Ascribed the Status of Being the First
- Behavioral Inferences
- Phylogenetic / Evolutionary Relationships
- Theories of Change
Introduction
The first fossil was found from Java in 1891 by the Dutch anatomist, Eugene Dubois. The exact locale was Trinil river bed in central java, and its common name assigned is “Java man”. The materials found were: a skull cap, a complete femur, three teeth, and a jaw fragment. This specimen was formerly referred to as “Pithecanthropus erectus” meaning an apeman who could walk straight. The femur found among the other materials showed prominent linea aspera, a criterion for erect posture and bipedalism while the skull cap denoted brain volume as that of present day apes. The dating of this fossil is middle pleistocene about 500,000 years ago
Later, in 1929 a Chinese paleontologist, W.C. Pei, discovered a dozen incomplete but identifiable skulls, mandibles, 147 teeth, and some fragments of the post-cranial skeleton. These also dated to middle pleistocene about 300,000 years ago. The exact locale was Zhoukoudian, some 30 miles southwest of Peking, China, which is why we assign the common name as “Peking man”. Its original name was “Sinanthropus pekinensis” meaning apeman from China.
It was later analyzed that the anatomical differences between these two fossil finds lie at the subspecific level. So, both the finds were placed under the same genus and species and are thus now known as: Homo erectus erectus and Homo erectus pekinensis respectively.
Several fossils that can be attributed to Homo erectus were later found from several sites of Europe and Africa. Here are some of the sites mentioned restricting to the most important and the best preserved specimens:
- Java
- Zhoukoudian and Lantien (China)
- Heidelberg – Germany
- Vertesszollos (Hungary)
- Arago (France)
- Ternifine (Algeria)
- Rabat (Morocco)
- Koobi Fora (Kenya)
- Olduvai gorge
- Swartkrans
- Saldanha
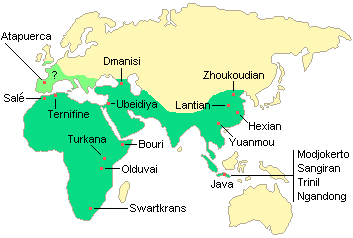
Dating the fossils
Evidence have led to the tentative conclusion that H. Erectus flourished over a long interval of pleistocene time. The fossils recovered at Koobi Fora are from about 1.7 mya, and Olduvai are probably 1.2 million years old. The specimens from Sangiran and Mojokerto in Java may approach the age of the Koobi Fora skeletons, and one from the Lantian localities in China is roughly contemporary with olduwai. The youngest hominins generally accepted as H. erectus are from Tighenif in Algeria (800–600 kya), Zhoukoudian in China (770–230 kya), and Sambungmacan and Ngandong (Solo) in Java (perhaps less than 250 kya).
Anatomy of Homo erectus:
- The cranial capacity of H. erectus averages about 1000 cc and generally ranges between 750 and 1250 cc.
- The size of the brain case of most of the specimens falls within the lower range of variation of modern Homo sapiens.
- The cranial bones are thick with thick brow ridges which are continuous forming a distinct supra orbital torus.
- There is a pronounced postorbital constriction.
- The skull is low and relatively flat, or platycephalic and in some specimens especially, in Java man, there is a bony ridge, the sagittal keel found along the midline at the top of the brain case.
- The profile of the cranium as seen from the side clearly shows: The angularity of the occipital, above this is a horizontal bar of bone, the occipital torus.
- The greatest width of the skull is towards the bottom. The facial skeleton is comparatively large and broad as compared to that of modern H. sapiens.
- The face, which is preserved in only a few specimens, is massively constructed, and its lower parts project forward.
- The bone forming the wall of the nose is thinner and the nasal bridge is relatively high and prominent.
- The teeth of H. erectus are smaller than the Australopithecus but larger than the H. sapiens. It is in accordance to the characteristics of the genus homo where teeth get reduced in size and number with time.
- The dental arcade is diverging with the greatest width occurring between the third molars.
- The mandible lacks chin, but does not have a mandibular torus.
- Externally the erectus femur resembles that of the sapiens but x-rays reveal that the outer wall of the shaft of erectus femur is twice as thick as that of the sapien’s femur.
- The presence of prominent linea aspera on the posterior side of the femur strongly suggests erect posture and bipedal locomotion for the H. erectus.
Cultural remains:
More than 100,000 artifacts have been recovered from the Zhoukoudian sites that were occupied intermittently for almost 250,000 years. According to Wu and Lin, 1983, it is one of the sites with the longest history of habitation by man or his ancestors. The occupation of the site has been divided into three cultural stages:
- Earliest stage (460,000-420,000 years ago) the tools are large, close to a pound in weight and made of soft stone such as sandstone.
- Middle stage (370,000-350,000 years ago) tools become smaller and lighter (under a pound) and these smaller tools comprise approximately two-thirds of the sample.
- Final stage (300,000-230,000 years ago) tools are still small, and the tool materials are of better quality. The coarse quartz of the earlier periods is replaced by finer quartz, sandstone tools have almost disappeared and flint tools increase in frequency by as much as 30 percent.
Expansion of the brain presumably enabled H. erectus to develop a more sophisticated tool kit than seen among earlier hominids. The important change in this tool kit was a core worked on both sides called bifaces (known widely as handaxe or cleaver). The biface had a flatter core than the roundish earlier Oldowan pebble tool. And probably even more important, this core tool was obviously a target design that is, the main goal of the tool maker. This greater focus and increased control enabled the stoneknapper to produce straighter edges resulting in a more efficient implement. This acheulian stone tool became standardized as the basic homo erectus all-purpose tool for more than a million years. It served to cut, scrape, pound, dig, and served as the most useful tool that has been found in Africa, parts of Asia, and later in Western Europe.
The early tools were crude and shapeless but become more refined over time. Common tools at the site are choppers and chopping tools, but retouched flakes were fashioned into scrapers, points, burins and awls.
Other aspects of culture
Stone was not the only material also used bone and probably horn. Found in the cave were antler fragments which had been hacked into pieces. Antler bases might have served as hammers and the sharp tines as digging sticks. Also found in abundance were deer skulls lacking facial bones as well as antlers were unearthed thus leaving only the brain cases intact. It can thus be inferred that they served as drinking bowls. The way of life of the H. erectus has traditionally been described as hunter- gatherers who killed deer and horses as well as other animals and gathered fruits, berries, and ostrich eggs. Fragments of charred ostrich eggshells, the abundant deposits of berry seeds were unearthed in the cave, and the flourishing plant growth surrounding the cave all suggest that meat was supplemented by the gathering of herbs, wild fruits, tubers and eggs. Layers of ash in the caves suggest the use of fire and the presence of hearths.
Scientists have unearthed more than 350 ancient tools in Konso, Ethiopia that were used by humans’ ancient ancestors. The tools, which span roughly 1 million years of evolution, show a gradual progression to more refined shaping.
Homo erectus thus has been ascribed as the firsts in-
- Making and using tools
- Making and using fire
- First cave dwellers
- First hunter-gatherers
- First to use animal hides for clothing
- First to migrate out of Africa
Dispersal of Homo erectus
The pleistocene which lasted more than 1.75 million years was a significant period in hominid evolutionary history and encompassed the appearance and disappearance of Homo erectus.
The life of the hunter-scavengers was nomadic and the woodland and savannah that covered the southern tier of Asia would have been an excellent environment for H. erectus. As the population grew, small groups budded off and moved on to find their own resource areas. This process repeated again and again led by Homo erectus out of the African continent to travel east to as far as Java as well as to the west to Europe.
Behavioral inferences:
At Zhoukoudian the remains of H. erectus were found in cave and fissure deposits. Although this does not prove that these hominids were habitual cave dwellers, the additional evidence of associated remains—such as stone, charred animal bones, collections of seeds, and what could be ancient hearths and charcoal—all points to H. erectus as having spent periods of time in the grottoes of Zhoukoudian. On the other hand, the remains of Lantian, Trinil, Sangiran, and Mojokerto, as well as Tighennif, Olduvai, and Koobi Fora, were all found in open sites, sometimes in stream gravels and clays, sometimes in river sandstones, and sometimes in lake beds. These suggest that H. erectus also lived in open encampments along the banks of streams or on the shores of lakes and also that proximity to water was crucial to survival. There is little doubt that mastery of fire was an important factor in colonizing cooler regions. Indeed, this discovery may have sped the migrations of ancient humans into the chilly, often glaciated expanses of prehistoric Europe. Sooner or later humans started cooking their food, thus reducing the work demanded of their teeth. This in turn may have played an important part in minimizing the evolutionary advantage of big teeth, since cooked food needs far less cutting, tearing, and grinding than does raw food. Nevertheless, burned bones are present, and these relics may still speak to the ability of the Zhoukoudian inhabitants to roast meat.
Other signs of the culture of H. erectus are implements found in the same deposits as their bones. Chopping tools and flakes made from split pebbles characterize both the Zhoukoudian and Dmanisi deposits; both are members of a so-called chopper chopping-tool family of industries. At Tighennif in northwestern Africa, H. erectus was found in association with totally different kinds of stone implements; these comprised double-edged hand axes and scrapers that have been characterized as representing what archaeologists call an early acheulian industry. This is part of the great acheulian hand-axe industrial complex, remnants of which are found widely spread over large parts of Europe and Africa. An acheulian industry is known also to emerge from Olduvai Gorge, is also known as the Oldowan industry. There is no sign that they buried their dead: no complete burials have been found, nor have graves, grave goods, or red ochre (a mineral used as a paint by later forms of hominins), either on or around any bones.
Phylogenetic / evolutionary relationships
A few researchers have generally opposed the view that H. erectus was the direct ancestor of later species, including homo-sapiens. Louis Leakey argued energetically that H. erectus populations, particularly in Africa, overlap in time with more advanced homo sapiens and therefore cannot be ancestral to the latter. Some support for Leakey’s point of view has come from analysis of anatomic characteristics exhibited by the fossils. By emphasizing a distinction between “primitive” and “derived” traits in the reconstruction of relationships between species, several paleontologists have attempted to show that h. Erectus does not make a suitable morphological ancestor for Homo sapiens.
Because the braincase is long, low, and thick-walled and presents a strong brow ridge, and they claim that H. erectus shows derived (or specialized) characteristics not shared with more modern humans. At the same time, it is noted, Homo sapiens does share some features, including a rounded, lightly built cranium, with earlier hominins such as H. habilis. For these reasons, some paleontologists (including Leakey) consider The more slender, or “gracile” H. habilis and H. rudolfensis to be more closely related to Homo sapiens than is H. erectus. If this much is agreed, there is still uncertainty as to how and where H. erectus eventually gave rise to Homo sapiens. Several general hypotheses have been advanced, but there is still no firm consensus regarding models of gradual change as opposed to scenarios of rapid evolution in which change in one region is followed by migration of the new populations into other areas.
Theories of gradual change
A traditional view held by some palaeontologists is that a species may be transformed gradually into a succeeding species. Such successive species in the evolutionary sequence are called chronospecies. The boundaries between chronospecies are almost impossible to determine by means of any objective anatomic or functional criteria; thus, all that is left is the guesswork of drawing a boundary at a moment in time. Such a chronological boundary may have to be drawn arbitrarily between the last survivors of H. erectus and the earliest members of a succeeding species (e.g., Homo sapiens).
Theories of punctuated change
This view suggests that species such as H. erectus may have exhibited little or no morphological change over long periods of time (evolutionary stasis) and that the transition from one species to a descendant form may have occurred relatively rapidly and in a restricted geographic area rather than on a worldwide basis. Whether any homo species, including our own, evolved gradually or rapidly has not yet been settled.