Chromosomal aberrations are broadly classified as numerical or structural aberrations.
Numerical Aberrations
Numerical aberrations are those that cause a change (addition or deletion) in the number of chromosomes. They are further classified as euploidy changes or aneuploidy changes. Euploidy is the condition when an organism gains or losses one or more complete set of chromosomes, thus causing change in the ploidy number. For example, triploid (3n), tetraploid (4n) etc. . Aneuploidy is the condition when an organism gains or losses one or more chromosomes and not the entire set. For example, trisomy (2n + 1), monosomy (2n – 1) . In humans, euploidy conditions do not exist because the extent of abnormality is too large to sustain life. Aneuploidy conditions, however, are more common and are manifested in disorders such as Down syndrome, Klinefelter syndrome and Turner syndrome
- Autosomal Trisomies : Trisomy is the condition where there is an additional copy of one chromosome. It is represented as 2n+1. Individuals, who are trisomics, thus show three copies of the chromosome rather than the normal two. It is usually observed that trisomies of the smaller chromosomes are more tolerated than trisomies of the larger chromosomes. This is expected, because additional copies of larger chromosomes contribute to larger genetic imbalance than additional small chromosomes. You will find it no surprise that the most common trisomy is of the shortest chromosome in human – chromosome 21. The other trisomies that have been reported include trisomy 13 (Patau syndrome) and trisomy 18 (Edward syndrome).
- Autosomal Monosomies : Autosomal monosomies have not been reported beyond birth in humans. Even the loss of the smallest chromosome is not compatible with life. Most known cases, therefore, are stillbirths and spontaneously aborted fetuses. It seems that loss of whole chromosomes cause too much genetic imbalance, which cannot support life. However, partial monosomies have been reported and well documented. Partial monosomy refers to the loss of a part of a chromosome while the rest of the chromosome is retained. Since such a partial monosomy is a chromosomal deletion.
- Allosomal Aberrations : Changes in number of allosomes (X and Y chromosomes in humans) are termed allosomal aberrations. The gain or loss of these chromosomes alters the phenotype leading to syndromes. For example, the loss of one X chromosome in females leads to turner syndrome (XO) and the excess of one X chromosome in males leads to Klinefelter syndrome (XXY). As mentioned earlier changes in allosomes cause changes in the primary and secondary sexual characters along with other manifestations. We shall look at the consequences of two such changes and the contrasting variation they produce.
Reasons for Numerical aberrations
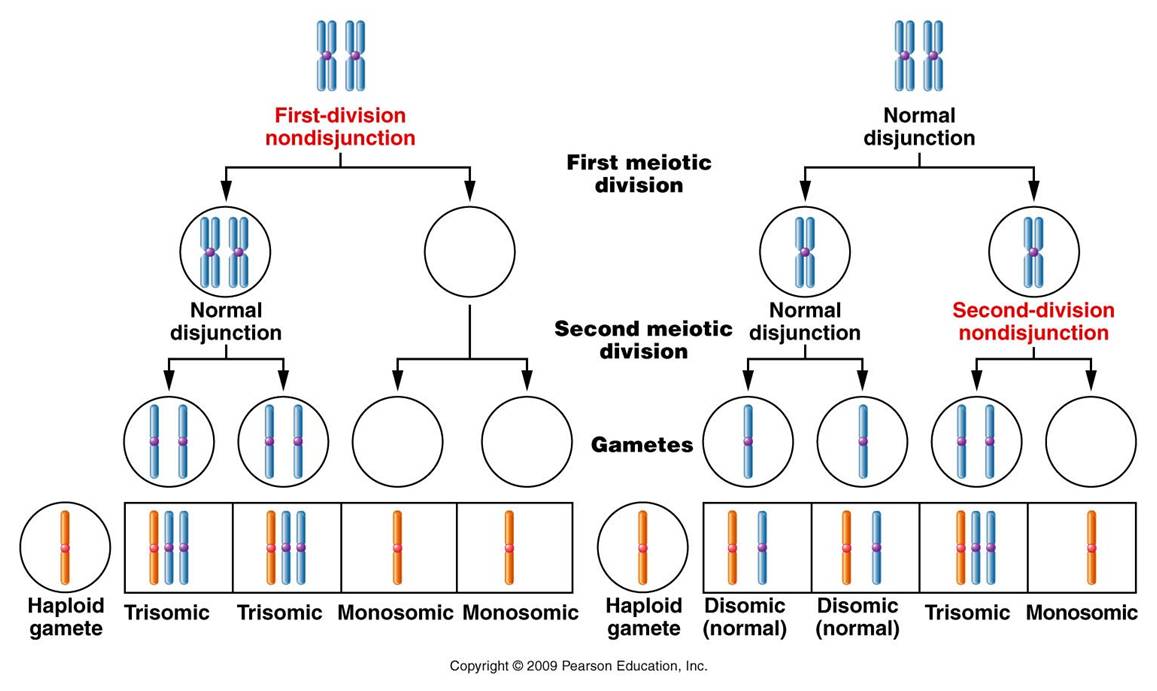
- Non-disjunction : It is the failure of separation of the chromosomes during mitosis or meiosis. Normal division involves the separation of the two arms (mitosis and meiosis-II) of the chromosomes or separation of the two chromosomes (meiosis-I) during the anaphase stage. This ensure that one copy of each is moved to each pole and consequently each daughter cell receives one copy. When this separation fails, both copies will move to one pole. Hence, one of the daughter cells will now have two copies while the other has no copies of that chromosome. Simply put, this is the basis of aneuploidy changes where there is one extra copy present or one copy missing in the cells. The normal meiotic and mitotic division and the consequences of non-disjunction at meiosis-I, meiosis-II and mitosis anaphase stages. The occurrence of non-disjunction is itself dependent on many factors. Some of these factors are:
- Advanced maternal age has been well correlated with an increase in the chances of non-disjunction. This is well illustrated in the fact that incidence of Down syndrome increases drastically as the maternal age increases. There is high correlation between high maternal age and incidence of Down syndrome (here Down syndrome is indicative of non-disjunction). This increase is attributed to the aging of the primary oocyte as age progresses and a reduction of the maternal competence to identify and abort abnormal fetuses. Increase in the time between ovulation and fertilization is well documented in animals to increase the rate of non-disjunction. As the frequency of copulations reduces there is an increased chance that there is a delay between ovulation and fertilization.
- Exposure to mutagens in general increases the chances of non-disjunction. Especially those who are constantly exposed to radiations have a high risk of non-disjunction.
- Genetic control of non-disjunction has been shown in a few species of Drosophila (fruit fly). These findings accounts for those few families that have shown to be prone to recurrent non-disjunction.
Structural Aberrations Chromosomal Aberrations
Structural aberrations are those that involve a change in the chromosome structure. These include deletions, duplications and rearrangements (inversions and translocations). Structural changes occur when chromosomes break and later rejoin in combinations that are different from the original. When there is a net loss or gain or chromosomal segments, the change is called an unbalanced structural change. When there is no net loss or gain of chromosomal segments, instead there is only a rearrangement; it is called a balanced structural change . Thus, balanced changes usually do not show any abnormal phenotypes, which unbalanced changes do. You should keep in mind that these changes are not mutations in genes; they only cause the number and order of genes to be changed. As with aneuploidy changes, structural changes are also seen in humans, and manifest in disorders such as Cri-du-chat syndrome, Wolf-Hirschhorn syndrome, Prader-Willi syndrome and Angelman syndrome. We shall discuss each of these changes and their effects later in the unit.
Reasons Structural Aberrations
(a) Robertsonian Translocation
As discussed before, Robertsonian translocation or centric fusion, causes a balanced rearrangement in the individual without any phenotypic abnormalities; however, due to improper meiotic segregation they give rise to trisomy-like and monosomy-like conditions in the offspring of such individuals.
A normal chromosomal complement in humans consists of two copies each of the 22 chromosomes and XY (for males) or XX (for females) – total of 46. Let us consider an individual who has a Robertsonian translocation between chromosomes 14 and 22. This person has a total of only 45 chromosomes. He
has two copies of all the other chromosomes except 14 and 21. For this pair of chromosomes he has one chromosome 14, one chromosome 21 and one translocation 14/21 chromosome.
In a normal individual, during meiosis, one copy of each of these chromosome moves to each pole. This results in daughter cells each containing one copy of 14 and one of 21. In a translocation individual, however, because there are three chromosomes instead of four two of them move to one pole and one moves to another pole. This causes abnormal chromosomal constituents in the daughter cells (gametes). There are different possible ways of these three chromosomes segregating . It is clear that only a small portion of gametes produced by individuals with such balanced translocations can produce normal offspring. Thus, a Robertsonian translocation can give rise to monosomies and trisomies of different chromosomes and their associated syndromes
Although these abnormalities do not cause true trisomies or monosomies, they give rise to conditions that are akin to true trisomies and monosomies. This is because, as stated before, the long arms of these chromosomes contain the bulk of the genes for that chromosome; presence of extra copies of the long arm has the same effect as having an extra copy of the entire chromosome.
(b) Reciprocal Translocation
Translocations not only cause trisomy-like and monosomy-like conditions, they also produce deletion-duplication conditions. A deletion-duplication is a condition where one segment of the chromosome is missing (deletion) and another is present in an extra copy (duplication).
Reciprocal translocations, wherein there is a mutual exchange of segments between two chromosomes, cause abnormal meiotic segregation. This abnormality is due to the formation of a quadrivalent of the four chromosomes during pairing . This structure is formed because the chromosomal segments always pair with their homologous regions. When such a complex structure is formed, separation of the chromosomes can happen in different ways depending on their orientation in the spindle. The different possibilities of segregation of a quadrivalent formed from reciprocally translocated chromosomes.
By analyzing the segregation products you should be able to predict the condition of the offspring from such a gamete. The first two segregation patterns produced phenotypically normal offspring. The next two segregation patterns may produce surviving offspring, but they will show abnormal phenotype due to the deletion duplication condition. Depending on the size of the del-dup segment the severity may vary. The last two segregation patterns are usually lethal. This is due to the del-dup segments being very large in these cases. If you recall, deletions of over 2% of the genome is incompatible with survival.
Hence, translocations by themselves do not cause deletions or duplications; it is only in the next generation that their effects are seen. It cannot be emphasized enough that a balanced translocation carrier will most probably have normal phenotype, unless the breakpoint disrupts some gene. As with all balanced rearrangements, we shall see that inversions, too, cause deletions and duplications because of abnormalities in meiotic division.
(c) Inversions
Inversions are balanced genetic rearrangements that invert segments within the chromosome. Depending on the involvement of the centromere they are either paracentric or pericentric . It is important to distinguish between these two types because the crossover products after meiosis is different for
each. Inversions too cause the formation of “inversion loops” during meiotic pairing. Because one of the two homologous chromosomes contains the inversion, it folds back into a loop to allow for maximum homologous pairing . Crossing over is a unique event in meiosis that causes recombination between the homologous pair of chromosomes. When crossing over occurs in a region within an inversion loop, it gives rise to recombinant products that contain deletion and duplication.
The formation of the inversion loop produces maximum homologous regions to be paired up. In pericentric inversions the inversion loop contains the centromere and in paracentric inversion the centromere is outside the inversion loop. Crossing over outside the inversion loop will give rise to normal chromosomes and inversion chromosomes. A crossover within the inversion loop, however, produces two non-recombinants (one normal and one inverted) and two recombinants (that contain deletion and duplication). These recombinants will contain duplication of certain genes along with deletion of other genes.
In pericentric inversions the deleted and duplicated segments do not involve the centromere; hence four types of gametes will be produced. Two of these will contain the aberrations; depending on the extent of the aberration it may or may not be compatible with survival. In paracentric inversions the deleted and
duplicated segments involve the centromere, hence we get one dicentric (containing two centromeres) and one acentric (containing no centromere) chromosome as recombinants. The dicentric chromosome forms a dicentric bridge during anaphase and thus arrests cell division (does not produce a gamete). The acentric chromosome is lost during division and thus doesn’t produce any viable gamete. Hence, only two types of gametes are produced from such individuals – one normal and one containing the inversion. These offspring will have normal phenotype because the inversion itself is a balanced rearrangement. Hence, the inversion itself will tend to persist in the population