INTRODUCTION
Children tend to resemble their patents in stature, body proportions, body composition, and rate of development. It may be assumed that, barring the action of obvious environmental determinants of growth, these resemblances reflect the influence of genes. that parents contribute to their biological offspring. Genes do not directly cause growth and development. Rather, the expression of a genetically inherited pattern of growth is regulated by many proteins that genes produce, and entire process is mediated by several biological systems, especially the endocrine and neurological systems.
GROWTH
The factors that affect the rate or tempo of growth must be dealt independently from factors that affect the size, shape and body composition of the child. The genetical control of tempo looks to be free of the genetical control of final adult size, and, to a great level, of final shape. Likewise, environmentally-produced alterations in tempo do not essentially affect final size or shape. Certainly, size and shape themselves seem to ~e independently controlled, by genetical and by environmental factors. The genetical mechanism of shape is too difficult than that of size, apparently because shape signifies primarily how the cells are distributed, while size characterises more the sum of sizes of the various cells. The number of cells is fixed early, in comparati ve safety of the uterus; their size continues to change during most of the childhood, and in some case’s, such as the fat ce]I, all through life.
(a) Genetics of Size
Monozygotic twins, who have the same genotype, usually resemble each other very closely indeed when brought up under similar circumstances. Table 1.1 shows the average differences in length or height from birth to four years between pain of’ monozygotic and of dizygotic same-sexed twins, from the Louisville data of Wilson, Dizygotic same-sexed twins resemble each other genetically no more closely than any other brothers or sisters, since they arise each from a different fertilized ovum, The monozygotic pairs were in fact less similar than the dizygotic at birth, but then this state quickly changed. Now this progressively close resemblance of the values in monozygotic twins echoes size, shape and tempo combined. Asymmetrical division of the ‘Original ovum, one twin getting just a little more cytoplasm than the other, and some what to their different positions in an overcrowded uterus could be responsible for differences between pairs at birth. The twin, who is smaller at birth, even if very little, ordinarily remains smaller throughout life. But if differ ence is large in size at birth, signifying some real intrauterine distress to the smaller twin, . then the differences later are quite large.
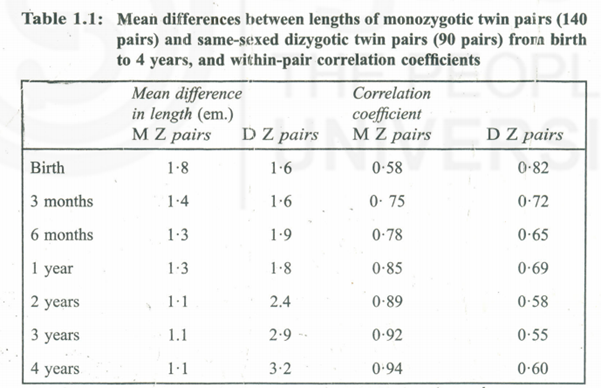
The extent to which height is under genetic control when environmental conditions are suitable is revealed in the variation within families compared with the variation amongst a population. Height is usually said to be controlled by many genes, each of small effect; such small-effect genes are termed ‘polygenes’ and are supposed to be positioned in parts of the chromosomes distinct from those carrying ‘major’ genes. The impression that height was under the influence of poly genes started because of the continuous and Gaussian distribution of height in the population. It is undisputable hat the summation of many small effects brings about precisely this type of continuous distribution but it is also reality that a distribution virtually indistinguishable from it may be conveyed out, by as few as five or s’ x major genes interacting. The question is thus an open one.
(b) Genetics of Shape
Some studies have shown that body measurements show higher correlations between parents and grown-up children than others. For instance, in a study of 125 Belgian families, comprising 282 grown-up children, Susanne (1975) found’ the correlations . In height each parent contributes equally to each offspring; despite all popula- belief to the contrary, there is r 0 tendency for daughters to resemble their mothers and sons their fathers significantly, Other data l:’l11y bear out this generalisation. Further down the table, the correlations for hip width closely resemble those for height. But the biacromial diameter ‘shoulder width) values are less similar. The father-son correlation is very low, and he father-daughter is lower than mother-daughter. Amongst the head measurements, lead breath resembles height in pattern though all the correlations are rather lower. but in head length the father’ correlation seems especially low, and in nose breadth not nears resemble one another not at all.
Arprenation of these correlations is not entirely straightforward. Measurements ch affected by environment ‘have low correlations; relatives simply do not amble one another very much. If a measurement is chiefly controlled by a few es IJf which one or more shows dominance (i.e., a single dose gives the same et as a double dose), the correlations’ decrease. The presence of sex-linked genes genes on the X chromosome), causes the sister-sister correlation to exceed the her-brother correlation; the father-daughter exceed the father-son; and the her-son to exceed the mother-daughter, No example of this pattern appears in table. What is clear is that different: meaurements show different degrees and patterns of familial resemblance. These differences cannot be understood until the development of each measurement is studied and the physiological factors, controlling its growth are clarified (Tanner, 1990).
Not all genes are active at birth. Some are not switched on till later, and the products of others can express themselves only in the physiological surroundings provided by the later years of growth. Some genes only produce their effect in one sex, usually because the gene product needs the co-operation of either one or th9 other hormonal environment in order to exert its action. These genes are called •sex-limited’ . The effect may not be all-or-none; the type of central baldness that is quite common in men depends on a gene which in single dose causes an effect in males only. In a double dose, which rarely occurs, it produces a similar effect in women (Tanner, 1990).
There is a difference between sex-limited genes and sex-linked genes-with sex-linked genes affecting alternately males and females in successive generations (so-called ‘criss-cross inheritance’). It has been observed that in sex-linkage the affected males are worse than the affected females. The reason being rnat the females have two X chromosomes and therefore two genes are in question, and one will be normal. Males have only one X chromosome and therefore lack the single normal gene. Hence, more males are affected in sex-linked disorders, as they have single X chromosome.
There are genes that are unable to express themselves till after birth, perhaps the resemblance in size between children and their parents is quite small during the first 12 to 18 months. During the first 18 months many babies change their centile positions for height and weight. Children with genes carrier for large size but born to small mothers move upwards through the centiles and children born large but with genes responsible for small size move downwards. T~,substantiate thisSmith et aI. (1976) quoted babies in a well nourished American middle-class population; the upward shift was generally complete within six months after birth, while the I downward shift took up to 18 months to realize. From age 2·0 till adolescence the parent-child correlation can be used as reference for children’s heights which provides precise means of diagnosing short stature than with the ordiJiary population standards. I The correlation coefficients between height measurements of a child at successive ages and his own measurement as a fully-grown adult describe the curve shown’ in Fig. 1.1. The correlation of length at birth with adult height is only about 0·3 since size at birth reflects uterine conditions much more than foetal genotype. Thereafter, the correlation rises steeply and by age three, it is of the order of 0·8. This means that the adult height can be predicted from height at age three, with an error which may amount to as much as 8 cm. either way. (It takes higher correlation than 0·8 to guarantee good prediction in the individual case). At puberty, the correlations diminish because of some children maturing early and some late, but if bone age is taken into account the prediction is restored (Tables of prediction of adult height from childhood height and bone age Tanner 1990). The correlation coefficients between height of the parents and height of the child at successive ages describe curves very similar to those shown in Fig. 1.1
(c) Tempo of Growth
The genetical control of tempo of growth (as contrasted to shape and size) is best witnessed by the inheritance of age at menarche. Monozygotic twin sisters who are growing up together under West European conditions attain menarche on average difference of two months while in dizygotic twins it is 12 months. Genetical influence is responsible for large section of the unpredictability in age at menarche. It is believed that mother and father exercise an equal influence on tempo of growth, so a late-maturing girl is as expected to have a late-maturing father as she is to have a late-maturing mother.
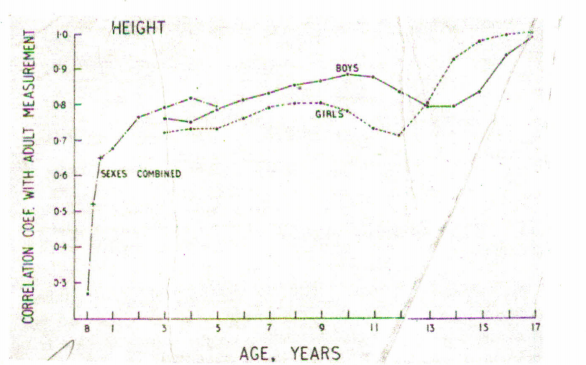
This control of tempo functions through the whole process of growth; the skeletal maturity shows the same type of family correlations as menarche at all ages. Similarly, the age of eruption of the teeth is controlled. In rational environmental circumstances the genetical control extends down to many of the details of the growth curves. From the records of three sisters shown in Fig. 1.2; on the left-hand figure the heights are plotted against chronological age and in the right-hand figure alongside developmental age, as given by years before and years after peak height velocity at adolescence. Note that two of the sisters have curves which are virtually super imposabJe apart from that they are on a different time-base, one being almost a year in advance of the other. So, the two differ drastically in one parameter of their growth curve, butlittle in their other parameters, although the third sister contrasts little from the other two in velocity (when plotted against developmental age the curves are ‘parallel ‘), but distinctly in absolute height.
(d) The Effects of Genetic Abnormalities on Growth
While, it is not an easy task to assign the hereditary and environmental contributions -to the phenotype, it is recognized that mutations to the DNA a person inherits, may perhaps produce various kinds of abnormal growth and development. Understanding what happens when the genetic material is disrupted, allows us to better understand the role of normal DNA in growth and development. Achondroplasia is a condition that results in short stature due to impaired growth of the legsand arms (there are other growth consequences as well). About 1115000 live births in Europeans are affected, making achondroplasia a frequent cause of skeletal, dwarfism. In 1994 a French research team announced the discovery of the gene defect responsible for ‘achondroplasia (Rousseau et al., 1994). The gene has been mapped to the short arm of chromosome four, and the DNA point mutation that alters the amino acid make-up of a protein blast growth factor receptor-J (Bonaventure et aI., I~96). The change in that receptor protein causes achondroplasia and several other related disorders (e.g., than atophoricdysplasia and hypochondroplasia). To date, 100 percent of patients with achondroplasia have the identical DNA mutation (Bogin, 1999).
Studies of people with unusual karyotypes (the number and type of chromosomes inherited by an individual) provide evidence for such genetic effects. Normal human karyotypes are 46, XY for males and 46, XX for females. A classic study by Tanner et al. (t959) examined people with sex chromosome anomalies, including individuals with 47, XXV (Klinefelter’s syndrome) and 45, X (Turner’s syndrome) karyotypes. People with the 47,XXY condition are phenotypically males and taller on average than normal 46, XY males. People with 45, X condition are phenotypically female, and much shorter, on ave!’ige than normal 46, XX females. Tanner et al. found that the body proportions (e.g., the ratio of leg length to stature) and rate of skeletal development 47, XXV boys was like that of normal 46, XY boys. They also found that the rate of skeletal development of 45, X girls, up to puberty, was like that of normal 46, XX girls. Consequently, the factors on the Y chromosome produce the male pattern of growth in body proportions and skeletal development.
An X chromosome effect on growth was proposed by Garn and Rohmann (1962). They used a longitudinal sample of hand-wrist radiographs and dental radiographs to study ossification rate (number of present), ossification timing (age at the appearance of a center), and tooth calcification in siblings. According to them the rates of skel tal and tooth development are genetically controlled and these genes are linked to the sex chromosomes. They also proposed that pairs of sisters, who share the same paternal X chromosome, should have greater concordance in rates of development than pairs of brothers, who have only a 50 percent chance of sharing the same maternal X chromosome, or brother-sister pairs, who share no paternal sex chromosomes. It was found that the correlation between pairs of sisters in skeletal and dental development (averaging about 0.52) was significantly greater than the correlation between pairs of brothers or bother-sister pairs (averaging about 0.35). Gam and Rohmann interpreted these correlations for X chromosome genetic control . for rates of development (Bogin, 1999).
The research strategy of searching for genetic determinant of growth by describing the size and development of individuals with sex anomalies has been used by many ,other investigators, including studies by Ratcliffe (1995). Ratcliffes work focuses on a longitudinal study of boys and girls with sex chromosome abnormalities who were identified at birth. In particular, her work concentrates on individuals with supernumerary sex chromosomes (XXX, XYY, XXV). She finds that chromosome has no detectable- effect on prenatal growth, as measured by weight, length, or head circumference at birth. In contrast, an extra X results in smaller size, and the XXX karyotype significantly reduces all three birth dimensions. During infancy and childhood, XYY and XXV boys grow faster than XY boys, and during adolescence the XYY boys have a greater peak height velocity than XY boys (Bogin, 1999).
THE ENDOCRINOLOGY OF GROWTH
Endocrine glands are pool of cells that manufacture chemical agents and release them into the blood-stream – the hormones. Strictly speaking, a hormone is any substance secreted by one cell which passes to and has an action on another. Some hormones act on cells very close to their origin and do not enter the blood; some enter the blood but only in a special portion of the circulation designed to carry them a short distance to their target; and some enter the general blood-stream and are carried in it all over the body. Thus- hormones constitute one of the two great systems of communication in the body (the nervous system being the other). This is as true for the carrying over of messages from one time to another as it is for the carrying of messages from place to place. The endocrine system is’ one of the chief agents for translating the instructions of the genes into the reality of the adult form, at the pace, and with the result, permitted by the available environment (Tanner, 1990). About dozen hormones are of particular importance in the control of human growth.
(a) Growth Hormone (GH)
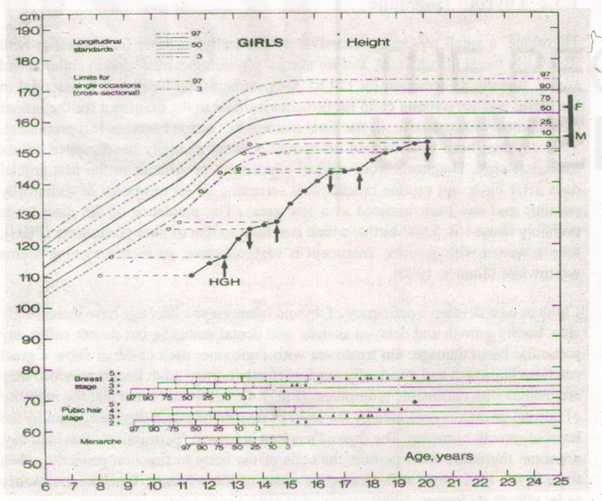
Growth hormone is essential for normal growth from birth to adulthood. Children deficient in it end up as adults about 130 cm. tall, but with normal proportions; they have been called miniatures. It is possible now to extract Growth hormone from humarr pituitary glands and administer to such children by injection; they then grow at a normal rate or even at faster rate than normal if small when the injections are started ..Fig. 1.3 shows the growth curve of a girl whose treatment started at 12·6 years.
Growth hormone is distinctly species-specific, although insulin and adrenocorticotrophic ACT H from animals work perfectly well in humans. Thereby it becomes essential that human growth hormone be extracted at autopsy because 191·amino acids OH is too large a molecule to be synthesized commercially, Luckily the pituitary glands of adults contain just as much OH as those of children.
OH is not required during foetal life, and its role in adults is unclear. Other than small size, adults with untreated OH deficiency are perfectly healthy and have no complaints. This could be because the secretion rate reduces after age 30 or so, regardless of the large quantities outstanding in the pituitary, where the concentration is 1,000 times that of any of the other pituitary hormones.
There is no continuous secretion of most pituitary hormones, but in a very rhythmic manner it bursts. OH comes under this. Majority of the time secretion of OH in the blood of children as well as adults are so low that it is scarcely detectable, but a few times in each 24 hours the levels escalates for periods of the order of 30 or 60 minutes. OH is habitually secreted some 60 to 90 minutes after sleep begins, and usually in response to exercise and anxiety. The mode of action of OH offers explanation for why merely episodic release is sufficient. Differences between normally large and normally small children and adults are perhaps due to receptors in the cartilage cells which control size. Anyway, normal small children have plenty of OH and they are not turned into normal big ones by being given OH in excess. In healthy children blood levels of somatomedin do not reduce with age to parallel the velocity curve, but, amazingly, increase from infancy to puberty. Thus, the normal velocity of growth depends on a more elaborate level of control.
(b) Thyroid Hormone
Thyroxine, a small molecule accessible synthetically, is basic for normal growth , from early foetal life onwards, and for normal physiological function in children and adults. It starts its secretion by 15th to 20th postmenstrual week and is essential in the foetus and very young child for protein synthesis in the brain and for the proper development of nerve cells. As the brain matures this action becomes less prominent. Thus children born with thyroid deficiency become mentally handicapped unless treated at once. Diagnosis from clinical appearance is difficult in the first crucial days after birth, but routine biochemical screening of all newboms is technically possible and has been initiated in a few areas. The incidence of the disorder is probably about 1 in 5,000 births, which is similar to that of phenylketonuria (PKU), long screened with success, Treatment is very effective, so such screening seems worthwhile (Tanner, 1990).
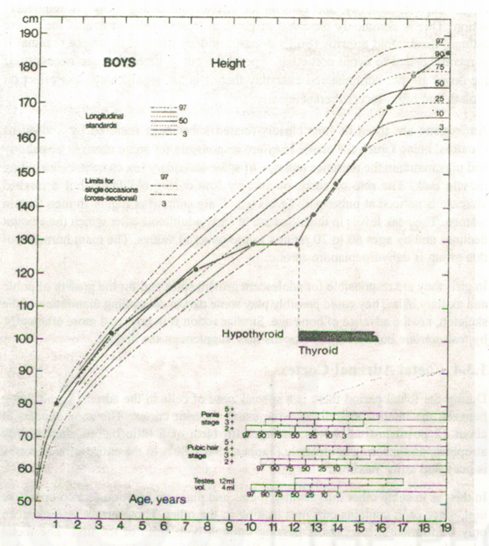
Children who develop a deficiency of thyroid hormone at a later age have increasingly slow bodily growth and delayed skeletal and dental maturity, but do not suffer any particular brain damage. On treatment with thyroxine, such children show a great catch-up in growth and eventually reach perfectly normal adult height provided they aretreated with reasonable promptness (Fig. lA). It does not seem that the intensity of normal growth is regulated by the level of thyroid hormone any more than by the level of growth hormone. The thyroid’s action is termed ‘permissive’; that is to say, a normal thyroxine level permits the cells of the body to function properly; when there is too little, most cells, including those of the pituitary, function too slowly to be effective (Tanner, 1990).
The rate of secretion of thyroxine is regulated by pituitary TSH (thyroid-stimulating hormone) itself controlled by hypothalamic Thyrotropin-releasing hormone (TRH). It is also called as Thyrotropin-releasing factor (TRF). Thyroxine level itself regulates TRH secretion, in a classical feedback loop.
(c) Adrenal Hormones
The adrenal gland comprises of an .inner core, the medulla, and an outer shell, the cortex. The two are distinct from a functional point of view, and the medulla, which secretes mostly adrenalin, contributing little to growth. The adrenal cortex secretes three groups of hormones:
Mineralocorticoids are aldosterone and to a smaller extent II-deoxycorticosterone. Aldosterone is necessary to life, but then has no direct bearing on growth.
Glueeeortlcolds (or corticoids) are cortisol (also caned hydrocortisone) and, to a lesser extent, corticoster one. Cortisol raises the formation of glucose from protein and has an anti-inflammatory and anti-stress action. Cortisol is secreted during foetal life, all through childhood and adults, with same quantity relative to body size. There is no particular increase at puberty anticipated from child’s increased size. Nevertheless, cortisol in normal amounts perhaps plays no role in controlling rate of growth, but if given in excess of normal, it slows up growth in height andretards skeletal maturity, and causing an increase in fat reflecting an antigrowth action. This is commonly observed in children treated with corticoids for severe asthma, rheumatoid arthritis (Still’s disease), kidney disease, or severe eczema. It is recommended to avoid corticoids if possible; but at times they are necessity. If the doses are not administered everyday then it is has significantly less effect on growth and often serves therapeutically.
Androgens are three or four closely-related substances secreted by androgens (‘andros’ being Greek for man). They are responsible for some changes of puberty and may maintain the presence and size of some secondary sex characters, including muscle bulk. The rate of secretion is very low during childhood but a marked increase is noticed at puberty, to levels which are somewhat higher in men than in women. The peak levels in the blood are in early adulthood; after which the amount declines and by ages 60 to 70 returns to pre-pubertal values. The main hormone of this group is dehydroepiandrosterone.
In girls they are responsible for adolescent growth spurt and for the growth of pubic and axillary hair. They could possibly play some part in controlling maturation of the skeleton, i.e. the advance of bone age. Similar action is in boys and more efficiently, by testosterone but adrenal androgens may supplement the action.
(d) Foetal Adrenal Cortex
During the foetal period there is a special zone of cells in the adrenal gland, lying between the interior medulla and the usual, exterior cortex. The zone appears at about 12 postmenstrual weeks, enlarges till birth or a little before, then swiftly atrophies, disappearing completely (Tanner, 1990). 80% of the entire adrenal cortex , is occupied at its maximum.
In this, as in some other interactions, foetus and placenta function as a co-operative unit, since each contains enzymes lacking in the, other. Thus certain materials can only be’ formed by precursors passing through both sides of the unit. The foetal adrenal cortex is a specialization peculiar to primates and does not occur in other mammals (Tanner, 1990).
(e) Testosterone
Testosterone a male sex hormone is secreted by the Leydig cells of the testis. There are three periods when level of testosterone rises in the blood. The first is in the foetus, where the rise begins at about 11 postmenstnial weeks· and lasts probably till birth. During this time the testosterone first causes differentiation of the external genitalia to form a penis and scrotum, and then sustains the further growth of the penis. It may also have an action on the brain, differentiating the hypothalamus irreversibly to the male, non-cyclic, type (Tanner, 1990).
Blood testosterone falls rapidly in the first few days after birth but a second period of high blood levels occurs during the subsequent six months, the peak being reached about two months after birth. By six months of age the level is down again to just detectable amounts and it continues thus till puberty. Then a very large increase occurs. At this time testosterone is responsible for growth of the penis, prostate and seminal vesicles, the pubic, axillary and facial hair, the growth of muscles and the adolescent spurt of the skeleton, especially the bones of the shoulders and vertebral column. The ‘presence of growth hormone is required for testosterone to exert its full effect on the muscles and on most of the skeleton. Since facial hair grows late and pubic hair early in the sequence, either the hair follicles at the two sites are unequally sensitive to differences in dose (pubic hair at low dose, facial hair only at high dose)or else there is a sequential maturation of testosterone receptors, this occurring earlier at the pubic site than at the facial. The latter is the more likely possibility, since when testosterone is given to a patient lacking it, pubic hair grow sat once, together with the penis and shoulders; but even at high doses the beard responds only much later. Testosterone secretion is controlled by pituitary LH (luteinizing or Leydig-cellstimulating hormone) (Tanner, 1990).
(f) Oestrogens
The ovaries secrete female sex hormones together known as oestrogens with oestradiol as main one. Its blood level is low till puberty. A large increase occurs then resulting in growth of the breasts, uterus and vagina, and development of the associated vaginal glands. It is also responsible for growth of parts of the pelvis. During menstruation oestradiol levels fluctuate regularly with the phase of the cycle, the secretion of Oestradiol is controlled by pituitary FSH (follicle-stimulating hormone).
(g) Gonadotropins
There are two pituitary gonadotrophins: follicle-stimulating hormone (FSH) and. luteinizing or Leydig-cell-stimulating hormone (LH) both secreting at low levels during childhood and increasing sharply at puberty. In males, ur is also secreted like GH, in a pulsatile fashion.
LH in females is secreted in a cyclic fashion and interacts with FSH to control the menstrual cycle. The egg grows under the influence of FSH and is shed; the cells immediately surrounding it remain behind in the uterus and form a body called the corpus luteum: LH stimulates this body to produce the hormone progesterone. Progesterone maintains the uterus in a state receptive to the implantation and growth of the ovum should it be fertilized; if no fertilization and implantation occurs, LH levels decline, the uterus sheds its lining in menstruation, and the cycle starts again (Tanner, 1990).
FSH in males causes growth of the seminiferous tubules, the sperm-producing parts of the testis. These occupy 90% of the volume of the testis, so the testicular enlargement at puberty is almost entirely due to them and not to the Leydig cells, however much the number of these increases.FSH is necessary for the growth of sperm, just as it is for the growth of the eggs. Much of the initial growth of eggs takes place during late foetal life, and in girls, though not in boys, FSH is especially high around 20-30 postrnenstrual weeks (Tanner, 1990).
(h) Prolactin
Prolactin, secreted by the pituitary is required for the secretion of breast milk in . the adult, but its role in endocrinology of childhood is unclear. The blood level in boys stays the same from early childhood onwards without any change at puberty, whereas in girls, the pre-pubertal level is the same as’ in boys, while small rise occurs in late puberty probably accountable for the filling out of the breast beneath the protuberant areola which causes the passage from the oestrogen-induced stage 4 to the adult stage 5.
(i) Insulin
Insulin is produced by the islets of Langerhans – assemblies of cells in the pancreas. The function of insulin causes glucose to absorb into cells, and stored as glycogen in the liver and muscles; deficiency of it results in sugar diabetes. It is present in both children and adults and has no specific action as far as growth is concerned except that its presence in normal amounts is required for normal growth to occur. Diabetic children whose sugar level is well controlled by injected insulin and suitable diet grow quite normally, but then a small degree of negligence in the control results in stunting and retardation of growth.
(j) The Endocrinology of Puberty
Though some information on mechanism of hormones producing change during puberty have been provided, yet the accurate details are not known. Although the methods to estimate the small amounts of hormones found in blood and urine are there but the very pulsatile nature of hormone release makes single blood assessment in many cases useless.
Adolescent growth spurt is result of joint actions of androgens and growth hormone. The 24-hour integrated blood levels of GH are higher in children than found in adults, but there appears no rise during the adolescent spurt. The standard level of GH must be present for testosterone to produce its full growth-effect on the muscles and on the bones of the limbs and shoulders although in the absence of GH the height spurt is only about two-thirds of normal, and the shoulder-width spurt even less. Interestingly enough the vertebral-column growth spurt in boys can apparently occur without GH. But in the absence of testosterone, no spurt takes place at all. In girls the height spurt attained is about two-thirds of normal in the absence’ of growth hormone; and the oestrogen induced growth of hip width is similarly reduced.
(k) Initiation of Puberty
The first episode in the categorization of puberty, straightaway preceding the morphological changes, is an increased secretion of the two gonadotrophic hormones of the pituitary caused by an increase in secretion of luteinizing hormone-releasing hormone (LHRH), also called as gonadotropin-releasing hormone (GnRH) by the, cells of the hypothalamus. Initially, the LHRH yields a comparatively small secretion of LH and FSH, may be because the LHRH receptors in the pituitary cells require some stimulation to sensitize’ them or the LH and FSH synthesizing cells need some practice. Progressively the response increases followed by rise in the blood levels of LH and FSH, and subsequently the peripheral events follow.
The stimulus to the secretion of LHRH in girls is as follows: a feedback system during childhood already exists with LHRH stimulating the production of FSH, FSH stimulating production of oestrogen, and oestrogen inhibiting the production of LHRH, The system operates, though, at a low level, and the amounts of oestrogen circulating are inadequate to stimulate breast or’ uterus growth. Then something occurs to render the hypothalamic sensor of oestrogen less sensitive. The low level of oestrogen is unsuccessful to stimulate it; LHRH is not inhibited and begins to rise. As a result, FSH rises, causing oestrogen to raise to a level enough to bring about the pubertal changes. Only at a point when high level of oestrogen ‘is reached is the sensor stimulated to turn off LHRH secretion. The feedback circuit is consequently re-established, however at a higher level.
Whatever could be the events that turns down the sensitivity of the oestrogen sensor, it characterises the culmination of a long chain of maturational happenings. Chronological age is not a good guide to the state of advancement while bone age is better as far as menarche is concerned. Marshall and Limongi (1976) demonstrated the use of bone age in conjunction with chronological age to predict menarche age. Nature and control of state of maturity are vague. It is expected to specify a hormonal cause of early and late maturation, and ,isolate and synthesize a tempo-hormone. Even bone age fails to predict the age of initiation of breast development or the first enlargement of testes. Children vary in how closely linked all their pubertal happenings are; in some the ‘clocks’ for breast growth and for menstruation run far apart, in others they are closely matched. The stimulus which alters the sensitivity of the oestrogen sensor comes from the nervous system.